后凹尾龙属查看源代码讨论查看历史
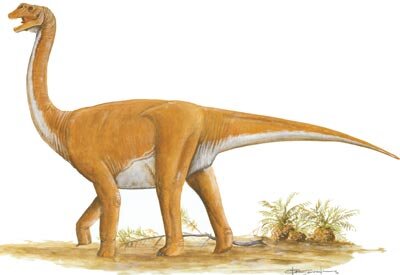 |
后凹尾龙属(属名:Opisthocoelicaudia),又名后空尾龙,是一属蜥脚下目恐龙,生存于白垩纪晚期,化石发现于蒙古的戈壁沙漠。
模式种是斯氏后凹尾龙(Opisthocoelicaudia skarzynskii)。1965年波兰和蒙古的科学家发掘出一具保存良好的骨骼,只缺乏头部和颈部,使后凹尾龙成为白垩纪晚期所知最详尽的蜥脚类之一。
骨骼上的齿痕可能代表著尸体曾被掠食者啃食,并带走了现在缺失的部位。迄今为止,只有额外两具不完整的标本,包含部分肩部及尾部碎片。后凹尾龙是种相对小型的蜥脚类,身长估计约11.4至13公尺。如同其他蜥脚类,他以长在长颈上的小头以及由四个圆柱状腿支撑著筒状身躯为特色。
后凹尾龙名字的意思为“后方空洞的尾巴”,代表著期不寻常的前段尾椎两侧向后方凹陷的情况。前述和其他骨骼特征使研究者推论后凹尾龙能够以后脚直立站起。
1977年由波兰古生物学家Maria Magdalena Borsuk-Białynicka命名及描述,后凹尾龙起初被认为是圆顶龙科的新物种,但现在的理论支持它是泰坦巨龙类的先进物种。后凹尾龙在泰坦巨龙类中的确切亲缘关系仍存在争议,它可能与北美洲的阿拉莫龙有接近亲缘关系。
所有后凹尾龙化石都来自耐梅盖特组,虽然当地有著丰富的恐龙化石,但该地层发现唯一的另一种蜥脚类是纳摩盖吐龙,所知来自单一头骨。鉴于后凹尾龙的头骨仍是未知,许多研究者提出纳摩盖吐龙和后凹尾龙可能代表著同一物种。耐梅盖特组的蜥脚类足迹包含皮肤印痕可能来自于两者之一,因为它们是当地所知唯二的蜥脚类。
发现与标本
模式标本发现于1965年6月10至23日之间,是一个由波兰古生物学家Zofia Kielan-Jaworowska所领导的波兰/蒙古的联合考察的期间。最大规模的一连串考察活动于1963年至1971年进行,共有21名成员参与以及更多雇用的蒙古工人协助。发现地点位于蒙古南部南戈壁省的Altan Uul地区,当地包含荒地约100平方公里。Altan Uul露出的沉积岩属于耐梅盖特组,是耐梅盖特盆地三个地层当中最年轻的一个。后凹尾龙是1965年考察活动众多重要恐龙中第一个发现的。其他发现位于不同地区,包含暴龙科特暴龙的多具骨骼、巨型似鸟龙类恐手龙的模式标本、蜥脚类纳摩盖吐龙、以及厚头龙类的平头龙。
在田野考察的第五天,考察团的地质学家Ryszard Gradziński发现了保存著良好骨骼的石块有望来自相当完整的骨骼。挖掘工作于隔天开始,揭露了几乎完整的骨骸只缺少头部和颈部。直到今日这具标本仍是这属恐龙发现最完整的标本。将标本运出崎岖地形引发了严重的技术问题,含著化石的石块必须在用汽油桶制成的简易金属雪橇上移动580公尺后才能装上卡车。因为骨骸埋在非常坚硬的砂岩层,许多岩块重达一吨。7月9日,骨骸包装成35个板条箱以运往达兰扎德嘎德。所有箱子包含石块及骨骸重达12吨。
模式标本属于一个老年个体。它的埋葬学方式不寻常,是背面倒地,其他耐梅盖特组的完整骨骸通常是腹面贴地。标本被发现包覆在交叉层理的河流沉积砂岩。大部分发现的脊椎仍然相连在一起,形成一系列包含8节背椎、6节荐椎、34节尾椎。另外3节脊椎被发现独立于系列外,可能属于颈部与背部相连的区域。骨骼的其馀部分略微偏离其原始解剖位置,左肢和左肋骨都被发现位于身体右侧,相反的右肢和右肋骨位于身体左侧。骨骸上以鉴定出咬痕,尤其在骨盆和大腿骨,代表著尸体曾经被掠食者啃食。头骨和颈部的失踪可能代表著这些部位被掠食者带走。遗骸的完整性代表著该个体就在发现地附近死亡。洪水事件可能将遗骸运输了很短的距离,然后甚至在软组织完全腐烂之前就被沉积物覆盖,。
1977年,波兰古生物学家Maria Magdalena Borsuk-Białynicka发表了对骨骼的全面描述并命名后凹尾龙为新属新种。属名指尾椎不寻常的两侧后凹情况,意为“后方空洞的尾巴”,由古希腊语的ὄπισθεν/opisthen(在后方)、κοῖλος/koilos(空心)以及拉丁语的cauda(尾巴)组成。
种名纪念处理模式标本的Wojciech Skarżyński。后凹尾龙是亚洲蜥脚类中,继盘足龙、马门溪龙后第三个得知颅后骨骼的。在出土后,模式标本成为华沙古生物机构的馆藏,但后来移交归还给来源国,现在保存于乌兰巴托蒙古科学院地质所,标本编号为MPC-D100/404。除了模式标本,Borsuk-Białynicka还叙述了来自同一地点的一副肩胛骨和鸟喙骨(ZPAL MgD-I/25c),这些骨头尚未彼此融合,推测来自青少年个体。
截至2017年,已从耐梅盖特组的32个地点发现蜥脚类骨骼,并可能属于后凹尾龙或纳摩盖吐龙。至少两具发现于耐梅盖特地区—一个破碎的尾部(MPD 100/406)和一对爪,展现了后凹尾龙的鉴定特征并可归其名下。由菲利浦·柯里领导的考察团于2006年和2008年尝试重新定位后凹尾龙模式标本的采集地,但直到2009年才成功,归功于Gradziński提供了额外的资讯。 尽管采集地已被风化填补无法找到额外的化石材料,但可确定该采集地来自耐梅盖特组下部层位。
叙述
如同其他蜥脚类,后凹尾龙有颗小头长在长颈上,一个筒状身躯由柱状四肢支撑,以及一条长尾。它是相对小型的蜥脚类,模式标本估计从头部到尾部尖端约11.4到13公尺。体重在不同的研究中分别估计为8.4吨、10.5吨、22吨、13吨、25.4吨[1]。
颅骨和颈部未保存下来,但颈韧带的重建表示有著中等长度约5公尺的颈部。在Borsuk-Białynicka的1977年叙述中指出具有11节背椎,葛瑞戈里·保罗则在2019年反驳已知的脊柱包含了第一节颈椎,而使具有泰坦巨龙类中典型的10节背椎。[2]就像其他泰坦巨龙类,因为缺乏椎骨副关节(Hyposphene-hypantrum articulations)而使背部非常有弹性,而骨盆则因附加的第六节荐椎而强韧。尾部的前段尾椎呈后凹型(opisthocoelous),意即前侧凸行、后侧凹型,形成杵臼关节。这种后凹型尾椎使后凹尾龙得名并将其独特于其他所有泰坦巨龙类。其他泰坦巨龙类通常以强度前凹型前段尾椎为特征,意即前侧凹入、后侧凸出。另一个独特特征可见于背椎,是双叉型神经棘突起,使沿脊柱顶部有两排骨质突出物。虽然这在泰坦巨龙类中很独特,却可以在其他不相关的蜥脚类中(如梁龙、盘足龙)发现,证明这些特征是独立演化的。
如同其他泰坦巨龙类的骨盆,坐骨相对较短,只有耻骨2/3的长度。左右坐骨及左右耻骨在大部分长度上都经过成骨作用,填补了其他蜥脚类这些骨头之间通常存在的缝隙。四肢比例上较短,也可见于其他泰坦巨龙类。前肢在几乎完整的标本内测量的高度为1.87公尺,大约是后肢长度的2/3,后肢被重建为高度2.46公尺就像其他泰坦巨龙类,四肢稍微向身体外侧扩展而非垂直于身体正下方,而前肢与其他蜥脚类相比更灵活及可动性佳。
手部仅由5个掌骨组成,呈垂直向并半圆形排列。腕骨消失,如同其他泰坦巨龙类。指骨和指爪也完全消失-在大部分泰坦巨龙类中,这些骨头依然存在但急遽缩小。在脚部,距骨像其他泰坦巨龙类一样大幅缩小,但跟骨完全消失。与手部相比,脚趾及趾爪发达。趾骨代数(phalangeal formula)代表从最内侧算起的趾骨个数为2-2-2-1-0。泰坦巨龙类的脚部骨骼非常罕见,除了后凹尾龙之外只有沉重龙和未命名因韦纳达河泰坦巨龙类发现保存完整的脚部,趾骨代数分别为前者2-2-3-2-0及后者2-2-2-2-0。在这三种泰坦巨龙类中,后凹尾龙最为进阶而具有最少的趾骨,表明在泰坦巨龙类的演化过程中趾骨数目的逐渐减少。与其他蜥脚类一样,第一趾爪不大于第二趾爪,但尺寸大致相等。
皮内成骨已在至少十种泰坦巨龙类发现。几乎完整的后凹尾龙骨骼缺乏皮内成骨可能代表其缺乏此构造。然而,亲缘关系接近的阿拉莫龙在首次发现后几乎过了一个世纪才发现具有皮内成骨,加上其他亲缘接近的泰坦巨龙类,使后凹尾龙具皮内成骨的可能性也增加。
参考文献
- ↑ Benson, R. B. J.; Campione, N. S. E.; Carrano, M. T.; Mannion, P. D.; Sullivan, C.; Upchurch, P.; Evans, D. C. Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage. PLoS Biology. 2014, 12 (5): e1001853. PMC 4011683. PMID 24802911. doi:10.1371/journal.pbio.1001853.
- ↑ Paul, Gregory S. Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals (PDF). Annals of the Carnegie Museum. 2019, 85 (4): 335–358.