抗體片段
| 抗體片段 |

{kind=link}
抗體(antibody)是指機體由於抗原的刺激而產生的具有保護作用的蛋白質。它(免疫球蛋白不僅僅只是抗體)是一種由漿細胞(效應B細胞)分泌,被免疫系統用來鑑別與中和外來物質如細菌、病毒等的大型Y形蛋白質,僅被發現存在於脊椎動物的血液等體液中,及其B細胞的細胞膜表面。抗體能識別特定外來物的一個獨特特徵,該外來目標被稱為抗原。
目錄
介紹
命名
抗體的結構
主要功能
特性和功能
多克隆單克隆
抗體的多樣性
抗體規律
介紹
抗體是一類能與抗原特異性結合的免疫球蛋白。抗體按其反應形式分為凝集素、沉降素、抗毒素、溶解素、調理素、中和抗體、補體結合抗體等。按抗體產生的來源分為正常抗體(天然抗體),如血型ABO型中的抗A和抗B的抗體,和免疫抗體如抗微生物的抗體。按反應抗原的來源分為異種抗體,異嗜性抗體,同種抗體和自身抗體。按抗原反應的凝集狀態分為完全抗體IgM和不完全抗體IgG等。
抗體在醫療實踐中應用甚為廣泛。如用於疾病的預防、診斷和治療方面都有一定的作用。臨床上用丙種球蛋白預防病毒性肝炎、麻疹、風疹等,國際上用抗Rh免疫球蛋白預防因Rh血型不合引起的溶血症。診斷上如類風濕因子用於類風濕性關節炎,抗核抗體(ANA)、抗DNA抗體用於系統性紅斑狼瘡,抗精子抗體用於原發性不孕症的診斷等;治療上如毒素中毒用抗毒治療以及免疫缺陷性疾病的治療等。
命名
19世紀後期,V on Behring及其同事Kitasato研究發現,用白喉或破傷風毒素免疫動物後可產生具有中和毒素作用的物質,稱之為抗毒素(antitoxin),隨後引入「抗體」一詞來泛指抗毒素類物質。抗體(antibody,Ab)是B細胞接受抗原刺激後增殖分化為漿細胞所產生的糖蛋白,主要存在於血清等體液中,是介導體液免疫的重要效應分子,能與相應抗原特異性結合,發揮免疫功能。
1937年,Tiselius和Kabat用電泳方法將血清蛋白分為白蛋白、α1、α2、β及γ球蛋白等組分,並發現抗體主要存在於γ區,因此抗體又被稱為γ球蛋白。隨後,經1968年和1972年的世界衛生組織和圈際免疫學會聯合會討論決定,將具有抗體活性或化學結構與抗體相似的球蛋白統一命名為免疫球蛋白(immunoglobulin,Ig)。Ig可分為分泌型Ig(secreted Ig,SIg)和膜型Ig(membrane Ig,mlg)。Slg主要存在於血液和組織液中,行使抗體的各種功能;mlg主要構成B細胞膜表面的抗原受體[2]。
抗體的結構
一、抗體的基本結構
經x線晶體衍射結構分析發現,Ig由四條多肽鏈組成,各肽鏈之間南數量不等的鏈間二硫鍵連接。Ig可形成「Y」字型結構,稱為Ig單體,是構成抗體的基本單位。
(一)重鏈和輕鏈
天然Ig分子含有四條異源性多肽鏈,其中,分子鼉較大的兩條鏈稱為重鏈(heavy chain,H),而分子量較小的兩條鏈稱為輕鏈(Light chain,L)。同一Ig分子中的兩條H鏈和兩條L鏈的氨基酸組成完全相同。
1.重鏈分子量為50 000~75 000,由450~550個氨基酸殘基組成。重鏈恆定區的氨基酸組成和排列順序不同,其抗原性也不同。據此,可將12分為5類(class),即IgM、IgD、IgG、IgA和IgE,其相應的重鏈分別為μ鏈、δ鏈、γ鏈、α鏈和ε鏈。不同類的Ig具有不同的特徵,如鏈內和鏈間二硫鍵的數量和位置、結構域的數量及鉸鏈區的長度等均不完全相同。即使是同一類的Ig,其鉸鏈區氨基酸組成和重鏈二硫鍵的數量、位置也不同,據此又可將同類Ig分為不同的亞類(subclass)。例如,人lgG可分為四個亞類,包括IgGl、IgG2、IgG3和IgG4;人IgA可分為IgAl和lgA2兩個亞類。[2]
2.輕鏈分子量約為25 000,由214個氨基酸殘基構成。輕鏈可分為兩種,分別為kappa(κ)鏈和lambda(λ)鏈。據此,可將lg分為兩型(type),即κ型和λ型。一個Ig分子上兩條輕鏈的型別總是相同的。不同類Ig既存在κ型,也存在λ型。同一個體內可同時存在κ型和λ型的Ig分子,不同種屬生物體內兩型輕鏈的比例不同。正常人血清Ig的κ:λ約為2:1,而在小鼠則為20:1。lg的κ與λ的比例異常可以反映免疫系統的異常。根據λ鏈恆定區個別氨基酸的差異,又可將λ鏈分為λl、λ2、λ3和λ4四個亞型(subtype)。
(二)可變區和恆定區
通過分析不同Ig重鏈和輕鏈的氨基酸序列發現,重鏈和輕鏈靠近N端的約1 10個氨基酸序列變化很大,其他部分氨基酸序列相對恆定。因此,將Ig輕鏈和重鏈中靠近N端氨基酸序列變化較大的區域稱為可變區(variable region,V),分別占重鏈和輕鏈的1/4和1/2;將靠近C端的氨基酸序列相對穩定的區域,稱為恆定區(constant region,C),分別占重鏈和輕鏈的3/4和1/2。
1.可變區 重鏈和輕鏈的V區分別稱為VH和VL。VH和VL中各含有3個氨基酸組成和排列順序高度可變的區域,稱為高變區(hypervariable region,HVR)或互補決定區(complementarity determining region,CDR), 包括HVRl(CDRl)、HVR2(CDR2) 和HVR3(CDR3),其中,HVR3(CDR3)變化程度更高。VH的3個高變區分別位於29~31、49~58和95~102位氨基酸,而VL的3個高變區分別位於28~35、49~56和91~98位氨基酸。VH和VL的3個CDR共同組成Ig的抗原結合部位(antigen-binding site),決定抗體的特異性,是抗體識別及結合抗原的部位。在V區中,CDR之外區域的氨基酸組成和排列順序相對保守,稱為骨架區(framework region,FR)。VH或VL各有四個骨架區,分別用 FR1、FR2、FR3和FR4表示。[2]
2.恆定區 重鏈和輕鏈的C區分別稱為CH和CL。不同型(κ或λ)Ig的CL長度基本一致,但是不同類Ig的CH長度不同,例如IgG、IgA和IgD包括CH1、CH2和CH3,而IgM和IgE則包括CHl、CH2、CH3和CH4。
(三)鉸鏈區
鉸鏈區(hinge region)位於CH1與CH2之間,富含脯氨酸,易伸展彎曲,從而改變抗原結合部位之間的距離,有利於抗體結合位於不同位置的抗原表位。鉸鏈區易被木瓜蛋白酶、胃蛋白酶等水解,產生不同的水解片段。不同類Ig的鉸鏈區不盡相同,例如人IgGl、IgG2、IgG4和IgA的鉸鏈區較短,IgG3和IgD的鉸鏈區較長,而IgM和IgE無鉸鏈區。
二、抗體的結構域
Ig分子的兩條重鏈和兩條輕鏈都可摺疊成數個球形結構域(domain),每個結構域行使其相應的功能。輕鏈有VL和CL兩個結構域;IgG、IgA和IgD的重鏈有VH、CH1、CH2和CH3四個結構域;IgM和IgE的重鏈有五個結構域,即多一個CH4結構域。每個結構域由約110個氨基酸組成,氨基酸序列具有相似性,其二級結構是由幾條多肽鏈摺疊形成的兩個反向平行的β片層(anti—parallel β sheet)構成的,兩個β片層中心的兩個半胱氨酸殘基由一個鏈內二硫鍵垂直連接,形成一個「β桶狀(βbarrel)」或「β-三明治 (β sandwich)」結構,這種折式稱為免疫球蛋白摺疊(immunoglobulin folding)。許多膜型和分泌型的蛋白質分子也含有這類獨特摺疊的二級結構,因此,這類分子被統稱為免疫球蛋白超家族(immunoglobulinsuperfamily,IgSF)[2]。
三、J鏈和分泌片
Ig輕鏈和重鏈除上述基本結構外,某些類別的Ig還含有其他輔助成分,如J鏈和分泌片。
(一) J鏈
J鏈(joining chain)是一條富含半胱氨酸的多肽鏈,由漿細胞合成,其主要功能是將多個Ig單體連接為多聚體。2個IgA單體由J鏈相互連接形成二聚體,5個IgM單體由二硫鍵相互連接,並通過二硫鍵與J鏈連接形成五聚體。IgG、IgD和IgE常為單體,無J鏈。
(二) 分泌片
分泌片(secretory piece,SP)又稱為分泌成分(secretory component,SC),是分泌型IgA分子上的一個輔助成分,為一種含糖的肽鏈,由黏膜上皮細胞合成和分泌,以非共價形式結合於IgA二聚體上,使其成為分泌型IgA(SIgA),並一起被分泌到黏膜表面。分泌片能保護SIgA的鉸鏈區不被蛋白水解酶降解。
四、抗體分子的水解片段
在一定條件下,Ig分子肽鏈的某些部分易被蛋白酶水解為不同片段。木瓜蛋白酶(papain)和胃蛋白酶(pepsin)是最常用的兩種Ig蛋白水解酶,並可籍此研究Ig的結構和功能,分離和純化特定的12多肽片段。
(一) 木瓜蛋白酶水解片段
木瓜蛋白酶水解Ig的部位是在鉸鏈區二硫鍵連接的兩條重鏈的近N端,可將Ig裂解為兩個完全相同的Fab段和一個Fc段。Fab段即抗原結合片段(fragment antigenbinding,Fab),由一條完整的輕鏈與重鏈的VH和CHl結構域組成。一個Fab片段為單價,可與抗原結合但不產生凝集反應或沉澱反應;Fc段即可結晶片段(fragment crystallizable,Fc),由Ig的CH2和CH3結構域組成。Fc段無抗原結合活性,是Ig與效應分子或細胞相互作用的部位。
(二)胃蛋白酶水解片段
胃蛋白酶作用於鉸鏈區二硫鍵所連接的兩條重鏈的近c端,水解Ig後可獲得一個F(ab』)2片段和一些小片段pFc 』。F(ab』)2是由兩個Fab段及鉸鏈區組成,由於Ig分子的兩個臂仍由二硫鍵連接,因此F(ab』)2片段為雙價,可同時結合兩個抗原表位,與抗原結合可發生凝集反應和沉澱反應。由於F(ab』)2片段既保留了結合相應抗原的生物學活性,又避免了Fc段免疫原性可能引起的副作用,因而被廣泛用於製備生物製品,如白喉抗毒素、破傷風抗毒素均是經胃蛋白酶消化後精製提純的生物製品。胃蛋白酶水解Ig後所產生的pFc 7最終被降解,無生物學作用。
主要功能
抗體的功能與其結構密切相關。同一抗體的V區和c區的氨基酸組成和順序的不同,決定了其功能上的差異。不同抗體的V區和C區在結構變化上具有一定的規律,又使得其在功能上存在共性。V區和C區的組成和結構,決定了抗體的生物學功能。
一、中和毒素和阻止病原體入侵
識別並特異性結合抗原是抗體的主要功能,執行該功能的結構是抗體的V區,其中CDR部位在識別和結合特異性抗原中起決定性作用。抗體有單體、二聚體和五聚體,因此結合抗原表位的數日也不相同。抗體結合抗原表位的個數稱為抗原結合價。Ig單體可結合2個抗原表位,為雙價。SIgA是二聚體,可結合4個抗原表位,為4價。IgM是五聚體,理論上可以結合10個抗原,應該是10價,但由於立體構象的空間位阻,使lgM一般只能結合5個抗原表位,故為5價。[2]
抗體的V區與抗原結合後,藉助於c區的作用,在體外可發生各種抗原抗體結合反應,有利於抗原或抗體的檢測和功能的判斷;在體內可中和毒素、阻斷病原體入侵、清除病原微生物;B細胞膜表面的IgM和IgD構成B細胞的抗原識別受體,能輔助B細胞特異性識別抗原分子。[2]
二、激活補體產生攻膜複合物使細胞溶解破壞
人IgG1~3和IgM與相應抗原結合後,可因構象改變而使其CH2和CH3結構域內的補體結合點暴露,從而通過經典途徑激活補體系統,產生多種效應功能,其中IgM、IgG1和IgG3激活補體系統的能力較強,IgG2較弱。IgA、IgE和IgG4本身難以激活補體,但在形成聚合物後可通過旁路途徑激活補體系統。通常情況下,lgD不能激活補體。
三、調理吞噬和ADCC
IgG可通過其Fc段與表面具有相應受體的細胞結合,產生不同的生物學作用。
1.調理作用(opsonization) 指IgG抗體(特別是IgG1和IgG3)的Fc段與中性粒細胞、巨噬細胞表面相應的Fc受體結合,從而增強吞噬細胞的吞噬作用。例如,細菌特異性的IgG抗體可通過其Fab段與相應的細菌抗原結合後,以其Fc段與巨噬細胞或中性粒細胞表面相應的Fc受體結合,通過IgG的Fab段和Fc段的「橋聯」作用,促進吞噬細胞對細菌的吞噬。
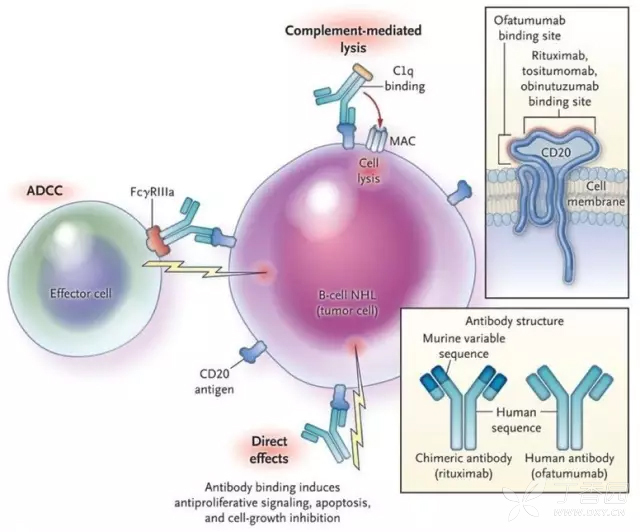
2.抗體依賴的細胞介導的細胞毒作用(antibody-dependent cell—mediated cytotoxicity,ADCC) 指具E有殺傷活性的細胞(如NK細胞)通過其表面的Fc受體識別包被於靶細胞表面抗原(如病毒感染細胞或腫瘤細胞)上的抗體的Fc段,直接殺傷靶細胞。 NK細胞是介導ADCC的主要細胞。抗體與靶細胞上的抗原結合是特異性的,而表達Fc受體細胞的殺傷作用是非特異性的。
四、介導 I 型超敏反應
IgE為親細胞抗體,可通過其Fc段與肥大細胞和嗜鹼性粒細胞表面的IgE高親和力Fc受體結合,使其致敏。當相同的變應原再次進入機體時,可以直接與致敏靶細胞表面的特異性IgE結合,促使這些細胞合成和釋放生物活性物質,引起I型超敏反應。
五、穿過胎盤屏障和黏膜
在人類,lgG是唯一能夠通過胎盤的抗體。胎盤母體一側的滋養層細胞可表達一種特異性的IgG輸送蛋白,稱為FcRn。IgG可選擇性地與FcRn結合,從而轉移到滋養層細胞內,並主動進入胎兒的血循環中。IgG穿過胎盤的作用在於這是一種重要的自然被動免疫機制,對於新生兒抗感染具有重要意義。另外,sigA可通過呼吸道和消化道的黏膜,在黏膜局部免疫中發揮重要的免疫防禦作用。
特性和功能
一、IgG
IgG於出生後3個月開始合成,3~5歲接近成人水平。IgG是血清和體液中含量最高的抗體,占血清總Ig的75%~80%。人lgG有4個亞類,根據其在血清中濃度的高低排序,分別為IgG1、IgG2、IgG3、IgG4。IgG的半衰期為20~23天,是再次免疫應答產生的主要抗體,其親和力高,在體內分布廣泛,具有重要的免疫效應,是機體抗感染的「主力軍」。IgG1、IgG2和IgG3可以穿過胎盤屏障,在新生兒抗感染免疫中起重要作用。IgG1、lgG2和IgG3能通過經典途徑活化補體,並可與巨噬細胞、NK細胞表面Fc受體結合,發揮調理作用、ADCC作用等;人IgGl、IgG2和IgG4可通過其Fc段與葡萄球菌蛋白A(SPA)結合,藉此可純化抗體,並用於免疫診斷。某些自身抗體如抗甲狀腺球蛋白抗體、抗核抗體,以及引起Ⅱ、Ⅲ型超敏反應的抗體也屬於IgG[2]。
二、IgM
IgM占血清Ig總量的5%~10%,血清濃度約為1mg/ml。單體IgM以膜結合型表達於B細胞表面,構成B細胞抗原受體,只表達mlgM是未成熟B細胞的標誌。分泌型IgM為五聚體,是分子量最大的Ig,沉降係數為19S,稱為巨球蛋白(macroglobulin),一般不能通過血管壁,主要存在於血液中。五聚體IgM含有10個Fab段,具有很強的抗原結合能力;含有5個Fc段,比IgG更易激活補體。天然血型抗體為IgM,血型不匹配的輸血,可導致嚴重的溶血反應。IgM是個體發育過程中最早合成和分泌的抗體,在胚胎髮育晚期的胎兒即能產生IgM,故臍帶血lgM升高提示胎兒有宮內感染(如風疹病毒或巨細胞病毒等感染)。IgM也是初次體液免疫應答中最早出現的抗體,是機體抗感染免疫的「先頭部隊」;血清中IgM升高,提示新近發生感染,可用於感染的早期診斷[2]。
三、IgA
IgA分為兩型:血清型為單體,主要存在於血清中,僅占血清Ig總量的10%~15%;分泌型IgA(secretory IgA,SIgA)為二聚體,由J鏈連接,含內皮細胞合成的分泌片,經分泌性上皮細胞分泌至外分泌液中。SIgA合成和分泌的部位在腸道、呼吸道、乳腺、唾液腺和淚腺,因此主要存在於胃腸道和支氣管分泌液、初乳、唾液和淚液。SIgA是外分泌液中主要的抗體類別,參與黏膜局部免疫,通過與相應病原微生物結合,阻止病原體黏附到細胞表面,在局部抗感染中發揮重要作用。SIgA在黏膜表面也有中和毒素的作用。新生兒易患呼吸道、胃腸道感染可能與IgA合成不足有關。嬰兒可從母親初乳中獲得SIgA,這是一種重要的自然被動免疫過程。
四、IgD[1]
正常人血清lgD濃度很低,儀占血清Ig總量的0.2%。IgD可在個體發育的任何時間產生。5類lg中,IgD的鉸鏈區最長,易被蛋白酶水解,故其半衰期很短(僅3天)。lgD分為兩型:血清型IgD的生物學功能尚不清楚;膜結合型IgD(mlgD)構成BCR,是B細胞分化發育成熟的標誌,未成熟B細胞僅表達mlgM,成熟B細胞可同時表達mlgM和mIgD,稱為初始B細胞(naive B cell)。活化的B細胞或記憶性B細胞表面的mlgD會逐漸消失。
五、IgE
IgE是正常人血清中含量最少的Ig,血清濃度極低,約為5×10-5mg/ml。IgE主要由黏膜下淋巴組織中的漿細胞分泌。其重要特徵為糖含量較高。IgE為親細胞抗體,其CH2和CH3結構域可與肥大細胞和嗜鹼性粒細胞上的IgE高親和力Fc受體結合,引起I型超敏反應。此外,IgE與機體的抗寄生蟲免疫相關。
多克隆單克隆
抗體獨特的生物學活性使其在疾病的診斷、免疫防治及基礎研究中發揮作重要作用。早在19世紀後期,人們就開始使用特異性抗原免疫動物製備相應的抗血清。1975年,Kohler和Milstein建立了單克隆抗體(monoclonai antibody,mAb)技術,使規模化製備高特異性、均質性抗體成為可能。然而,鼠源性mAb在人體反覆免疫後出現的人抗鼠抗體(human anti—mouseantibody,HAMA)很大程度上限制了mAb的臨床應用。近年來,隨着分子生物學的發展,人們已經可以通過抗體丁:程技術製備人一鼠嵌合抗體、人源化抗體或人源抗體。
多克隆抗體
天然的抗原分子中常含有多種不同的抗原表位,以該抗原刺激機體的免疫系統可同時激活多種B細胞克隆,產生的抗體中會含有多種針對不同抗原表位的抗體,因此稱之為多克隆抗體。多克隆抗體主要從動物免疫血清、恢復期患者血清或免疫接種人群的血清中獲得。多克隆抗體的優勢是:作用全面,具有中和抗原、免疫調理、補體依賴的細胞毒作用(CDC)、ADCC等重要作用,而且來源廣泛、製備簡單。其缺點是:特異性不高、易發生交叉反應,不易大量製備,因而限制了其應用的範圍。
單克隆抗體
解決多克隆抗體特異性不高的理想方法是製備識別單一表位特異性的抗體。如果能獲得僅針對單一表位的漿細胞克隆,並使其在體外擴增分泌抗體,就有可能獲得單一表位特異性的抗體。然而,漿細胞在體外的壽命較短,難以培養。為克服這一缺點,Kohler和Milstein將可產生特異性抗體但短壽的B細胞與不產生抗體但長壽的骨髓瘤細胞融合,獲得了可以產生單克隆抗體的雜交瘤細胞,從而建立了單克隆抗體製備技術。通過該技術融合形成的雜交瘤(hybridoma),既具有骨髓瘤細胞大量擴增和永生的特性,又具有免疫B細胞合成和分泌特異性抗體的能力。
每個雜交瘤細胞由一個B細胞融合而成,而每個B細胞克隆僅識別一種抗原表位,因此經篩選和克隆化的雜交瘤細胞僅能合成和分泌識別單一抗原表位的特異性抗體,稱為單克隆抗體。其優點是結構均一、純度高、特異性強、效價高、血清交叉反應少、製備成本低;缺點是鼠源性mAb對人具有較強的免疫原性,反覆免疫人體後可誘導產生人抗鼠抗體,從而削弱了其作用,甚至導致機體組織細胞的免疫病理損傷,因此需要進一步通過抗體工程技術製備人一鼠嵌合抗體、人源化抗體或人源抗體。
單克隆抗體(monoclonal antibody,Mab)技術是20世紀免疫學技術的一項里程碑式突破.該技術將免疫小鼠的B淋巴細胞與小鼠骨髓瘤細胞融合生成雜交瘤細胞,這種雜交瘤細胞核內含有雙親細胞的染色體,繼承了親代細胞的特徵.它既具有瘤細胞在體外培養中迅速增殖的能力.又具備免疫脾細胞合成和分泌特異性抗體的特性。
隨後用適當方法把雜交瘤細胞分離出來,進行單個細胞培養,使之大量繁殖,在培養液中形成單個雜交瘤細胞的克隆(也稱細胞系)。由於每個B淋巴細胞只有合成一種抗體的遺傳基因,所以單個雜交腐細胞的克隆也只能產生一種專一性抗體,即單克隆抗體。這種製備產生單克隆抗體的技術被稱為單克隆抗體技術。
由雜交瘤單細胞克隆所產生的單克隆抗體只能特異性地與抗原分子上的一個抗原決定簇結合,抗體成分均一,抗體的結構、氨基酸順序、特異性等都是一致的,且在培養過程中只要不發生變異,不同時間內分泌的抗體都能保持同樣的結構和功能。用這種技術可按需要生產大量很純的單一抗體,這些是用普通血清學方法所不能達到的。
單克隆抗體的發展經歷了鼠源性單克隆抗體、嵌合性單克隆抗體、人源化單克隆抗體和全人源單克隆抗體四個階段。特別是全人源單克隆抗體,其可變區和恆定區都是人源的,這類抗體藥物具有高親和力、高特異性、幾乎沒有毒副作用等優點,克服了動物源抗體及嵌合抗體的各種缺點,成為治療性抗體藥物發展的必然趨勢。
單克隆抗體技術在臨床應用中為疾病的診斷、治療提供了新手段,作為治療用藥物,單克隆抗體主要應用於腫瘤、自身免疫疾病、器官移植排斥及病毒感染等領域。單克隆抗體也可用於腫瘤的導向治療,將針對某一腫瘤抗原的單克隆抗體與化療或放療藥物連接,利用單克隆抗體的專一性識別結合特點,將藥物攜帶至靶細胞並直接將其殺傷。由於單克隆抗體具有特異性強、純度高、均一性好等優點,大大促進了單克隆抗體檢測試劑盒的發展,在病原微生物、腫瘤、免疫細胞、激素及細胞因子的檢測診斷中廣泛應用。若將放射性標記物與單克隆抗體連接,注入患者體內後可進行放射免疫顯像,協助腫瘤的診斷。在親和色譜中單克隆抗體是重要的配體,若將單克隆抗體固定到一個惰性的固相基質上,則可用於特異性抗原分子的高度純化[3]。
抗體的多樣性
人血清中的抗體多種多樣,B淋巴細胞可產生的抗體種類在108以上,可與眾多不同抗原發生特異性結合。抗體多樣性的原因主要有兩方面:
1.外源性因素環境中抗原種類甚多,每種大分子抗原又有多種抗原表位,每種抗原表位均可選擇激活體內一個B細胞克隆,產生一種特異性抗體。
2.內源性因素抗體多樣性的另一個原因是由基因的結構及功能特徵所決定的。編碼人Ig重鏈及κ、λ型輕鏈的基因分別位於第14、2、22號染色體上。其中編碼Ig重鏈的基因包括編碼可變區的V、D、J及編碼恆定區的C基因;編碼Ig輕鏈的基因包括編碼可變區的V、J及編碼恆定區的C基因。每種基因片段是以多拷貝的形式存在,其中編碼重鏈V區的VH、DH和JH的基因片段數分別為50、23和6個;編碼K輕鏈V區的Vx和JK基因片段數分別為60和5個,編碼入輕鏈V區的VX和J入基因片段數分別為30和7個。這些基因在胚系階段以分隔的形式存在。在B細胞的分化發育過程中,這些基因片段發生重排和組合,從而產生數量巨大、能識別特異性抗原的BCR。每種具有特異性BCR的B細胞克隆可識別相應的抗原,產生一種特異性抗體。Ig基因重組是B細胞合成無數特異性抗體的主要原因。
抗體規律
凡能產生抗體的高等動物(包括人類),當注入胸腺依賴性抗原(TD抗原)進行免疫時都有着相同產生抗體的規律,即存在初次免疫應答(primary immune response)和再次免疫應答(secondary immune response)。初次免疫應答是指機體第一次接觸某種抗原物質引起特異性抗體產生的過程。其特點是潛伏期長(一周以上),產生的抗體滴度(效價)低、維持的時間短,產生的抗體以IgM為主;再次免疫應答是指機體以後再次接觸同樣的抗原後所產生的抗體應答過程。其特點是產生抗體的潛伏期短、抗體滴度高,維持的時問長,產生的抗體以IgG為主。
非胸腺依賴性抗原(TI抗原)引起的體液免疫由於不產生記憶細胞,因此只有初次免疫應答,沒有再次免疫應答。