細胞核查看源代码讨论查看历史
 |
细胞核(nucleus}})是存在於真核細胞中的封閉式膜狀细胞器,內部含有細胞中大多數的遺傳物質,也就是DNA。這些DNA與多種蛋白質(如組織蛋白)複合形成染色質。而染色質在細胞分裂時,會濃縮形成染色體,其中所含的所有基因合稱為核基因組。細胞核的作用,是維持基因的完整性,並藉由調節基因表現來影響細胞活動。
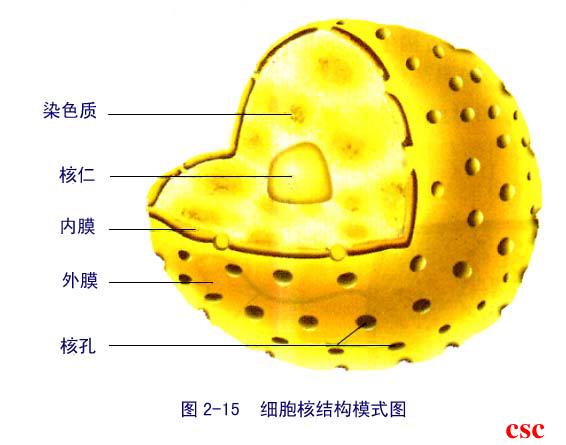
細胞核的主要構造為核膜[1] ,是一種將細胞核完全包覆的雙層膜,可使膜內物質與細胞質、以及具有細胞骨架功能的網狀結構核纖層分隔開來。由於多數分子無法直接穿透核膜,因此需要核孔作為物質的進出通道。這些孔洞可讓小分子與自由通透;而如蛋白質般較大的分子,則需要攜帶蛋白的幫助才能通過。核運輸是細胞中最重要的功能;基因表現與染色體的保存,皆有賴於核孔上所進行的輸送作用。
細胞核內不含有任何其他膜狀的結構,但也並非完全均勻,其中存在許多由特殊蛋白質、RNA以及DNA所複合而成的次核體。而其中受理解最透徹的是核仁,此結構主要參與核糖體的組成。核糖體在核仁中產出之前,會進入細胞質進行mRNA的轉譯。
目录
結構
細胞核對動物而言是最大的胞器。哺乳類細胞核的平均直徑一般為11到12微米(μm),佔據了細胞中大約10%的體積。細胞核內部的黏液稱為核質,與核外的細胞質類似。
核膜與核孔
核膜包括以平行方式相互重疊的兩層膜狀構造,也就是內膜及外膜,兩者之間的距離約10到50奈米(nm)。核膜將細胞核完全包覆,使內側的遺傳物質與外側的細胞質分離。並阻擋大分子在核質與細胞質之間自由擴散。細胞核的外膜與另一種膜狀構造粗糙內質網相連,兩者皆綴有核糖體。內外膜之間的空間稱為核膜間隙,這些空間與粗糙內質網中的內腔相連。
穿透核膜的核孔擁有類似於通道的功能,是由多種核稱為核孔蛋白的蛋白質所組成。核孔的分子量約125百萬Da,含有約50(酵母菌)到100(脊椎動物)個蛋白質。核孔的直徑為100奈米,不過真正可讓分子自由擴散的孔道只有寬9奈米,這是因為核孔中間存在一些調節系統。小型的水溶性分子可以直接通過,而大型分子如核酸與蛋白質則會受到阻礙,需要透過主動運輸才能進入細胞核。典型的哺乳類細胞核膜上,擁有約3000到4000個核孔。伴隨這些環狀構造的核籃(nuclear basket),則向內延伸進入核質;另外還有一系列的絲狀構造伸入細胞質中。這些構造的功能是用來與核運輸蛋白結合。
許多蛋白質、核糖體次單元或RNA,可在一類稱為核轉運蛋白(karyopherin)的運輸因子中介下通過核孔。其中可幫助分子進入核內的又稱為內輸蛋白(importin);幫助分子離開細胞核的則稱作外輸蛋白(exportin)。大多數核轉運蛋白可直接與欲運送的分子作用,有些則需要轉接蛋白協助。類固醇激素如皮質醇與醛固酮,以及其他作為細胞信號的脂溶性分子,可以從細胞膜擴散進入細胞質,並與將要進入細胞核內的核受體蛋白結合。這些受體與配體結合時具有轉錄因子的功用,若配體不存在,受體則有組織蛋白去乙醯酶的作用,可抑制基因表現。
細胞骨架
動物細胞內有兩種用來支撐細胞核的中間纖維:其中核纖層為一種有系統的網狀結構,分佈於核模內側;而另一種較缺乏系統的支撐構造則位於核模外側。兩種結構除了支撐核模外,也是染色體與核孔的賴以固定的位點。
核纖層主要是由層蛋白所構成,與多數蛋白質相同,層蛋白是合成於細胞質,之後再送入細胞核內部。這些蛋白質在核內會先聚集在一起,再與原有的核纖層網狀構造結合。此外,層蛋白也會出現在核質內部,組成另一種可在螢光顯微下觀察,稱為nucleoplasmic veil的調控結構。此構造位於核仁外側,且存在於分裂間期,其功能則尚未明瞭。目前已知有些形成veil的層蛋白結構,會與染色質結合並破壞其構造,進而抑制蛋白質編碼基因的轉錄。
與其他中間纖維相同的是,層蛋白單體含有一個α螺旋結構域。這些結構域兩兩互相纏繞,形成一種稱為捲曲螺旋的雙體結構。而兩個雙體還會再以反平行方式,組合成一種稱為原絲的四聚體。八條原絲可以在水平排列下,形成捲曲狀的繩狀纖維。這些纖維可以在相同狀態下聚合或分解,因此纖維的長度,是取決於纖維的增加與減少速率之間的競爭。
當層蛋白基因發生突變時,會導致纖維的聚合情形發生缺陷,此種狀況稱為層蛋白病。這類病症中,以一系列稱為早衰症的疾病較為著名。罹患早衰症的人,會顯現出提早成熟並老化的現象。至於基因與老化表型之間在生物化學上的詳細機制,目前並不明瞭。
染色质
染色质主要由蛋白质遗传物质DNA和构成,可以被碱性染料染成深色,在细胞分裂的间期,染色质呈现细长丝状;在分裂期,它们们会缩短变粗成为染色体,此时是观察染色体的最佳时期。此外还有少量遗传物质存在于线粒体、叶绿体等细胞器中。
染色質可分為兩種,一種是DNA以較鬆散的方式組成的真染色質,其中含有細胞中較多可表現的基因。另一種則是DNA結構較為緊密的異染色質,其中的DNA鮮少被轉錄。異染色質又可分為兩種,一種稱為「選擇性」異染色質,含有的基因在特定種類細胞或特定發育階段才會表現;另一種稱為「永久性」異染色質,內含一些染色體構成物,例如端粒或著絲粒。染色質在分裂間期會分別組織在各自的領域中,這些領域稱為「染色體區域」。主要存在於真染色質內的可作用基因,傾向於靠近在染色質區域的邊緣地帶。
可與特定染色質結構,尤其是與核小體結合的一些抗體,與一些自體免疫疾病,如全身性紅斑狼瘡有關。這些抗體稱為抗核抗體(ANA),已知與一部分發生於多發性硬化症中的全身性免疫系統失調有關。在早衰症中,抗體在導致自體免疫疾病症狀上的影響並不顯著。
核仁
核仁是細胞核內部一種染色濃度較高,且非膜狀的不連續構造,有時被稱作「次胞器」(suborganelle)。這些核仁是從rDNA,也就是核糖體RNA(rRNA)的DNA編碼周圍開始成型,rDNA片段具有串聯重複特性,稱作核仁組織區域(nucleolar organizer regions;NOR)。核仁的主要作用,是合成rRNA並組成核糖體;而核仁的凝聚性,則視其活性而定。當核糖體進行合成時,組成核仁的原料會快速聚合在一起,以幫助核糖體的生成,而核仁也並因此成型。觀察顯示當rDNA失去作用時,會使核仁的結構混雜在一起,使上述模型獲得支持。
核糖體組成的第一個步驟,是rDNA的轉錄,參與此過程的酵素是RNA聚合酶I。轉錄作用會生成rRNA前體,之後再被切割成三個次單元,分別是5.8S、18S以及28SrRNA。核仁中的rRNA會在轉錄以及後轉錄過程中聚集在一起,形成小核仁RNA(snoRNA)分子,其部分結構是來自被剪接作用移出的內含子,這些內含子原本屬於mRNA前體;而此過程裡的mRNA,則是由負責核糖體功能的基因所轉錄而成。組裝完成的核糖體次單元,是各種進出核孔的分子中體積最大者。
在電子顯微鏡的觀察下,可見核仁含有三個不同的區域,其中位於最內部的稱為「纖維中心」(fibrillar centers,FCs),往外一層為「緻密纖維組分」(dense fibrillar component,DFC),最外圍則是「顆粒組分」(granular component,GC)。rDNA的轉錄可發生於FC或是FC與DFC的邊界上。因此當細胞中的rDNA正在轉錄時,會使FC的數量增加。而大多數rRNA的分割與修飾,則發生在DFC部分,之後再於GC部位與蛋白質會合。
機能
細胞核的主要機能是調控基因表現,並調節細胞週期中的DNA複製過程。細胞核是轉錄作用所發生的位置,由與可與細胞質中的轉譯作用隔離,使真核生物得以擁有一些原核生物所缺乏的基因調節能力。
細胞分隔
核膜使細胞核可掌控其內容物,並使這些物質與外部的細胞質隔離開來。這種作用對於核膜內外兩側的各種調控處理作用皆相當重要。有時細胞質中的處理作用必須受到限制,這時參與作用的關鍵物質就會移動到細胞核中,並與轉錄因子作用以進行負向調節,進而抑制反應路徑中特定酵素的生成。以降解葡萄糖並生成能量的反應途徑糖解作用為例,參與其中的己糖激酶負責催化第一個步驟,使葡萄糖轉變成葡萄糖-6-磷酸。當糖解作用的後期產物果糖-6-磷酸濃度較高時,調控蛋白就會將六碳糖激酶移入細胞核。
為了調控正在轉錄當中的基因,細胞可將某些參與調節基因表現的轉錄因子與DNA隔離,直到這些因子在其他訊息傳遞路徑中活化。不必要的基因表現即使只有少量,也會在此過程中受到阻礙。以參與大部分發炎反應的NF-κB調控基因為例,轉錄作用會受一連串訊息路徑所誘導,一開始是TNF-α訊號分子與細胞膜上的受器結合,導致訊號蛋白的重新補充,最後使NF-κB受到活化。NF-κB上的核定位信號可使其經由核孔運輸進入細胞核內,刺激目標基因的轉錄。
分隔作用也可以防止細胞對尚未經過剪接的mRNA進行轉譯,因為真核生物的mRNA必須在轉譯作用開始之前去除其中的內含子,才能合成有用的蛋白質。mRNA在與核糖體接觸並開始轉譯以前,會先在細胞核中進行剪接作用,如果沒有細胞核,那麼未經處裡的mRNA就會受到轉譯,進而產生形狀錯誤,且功能無用的蛋白質。
基因表現
基因表現的第一步是轉錄,此過程利用DNA作為模板來合成RNA。具有蛋白質編碼的基因,會轉錄生成信使RNA(mRNA),而mRNA則會在稍後的轉譯作用中,經由核糖體的作用合成出蛋白質。由於核糖體位於細胞核外側,因此mRNA必須在合成後送出核外。
因為細胞核是轉錄進行位置,所以含有多種不同的蛋白質,有些直接參與轉錄;有些則參與相關的調節作用。這些蛋白質包括可以將雙股DNA螺旋解開,幫助其他蛋白與其接觸的螺旋酶;或是可以合成RNA的RNA聚合酶;以及可以改變DNA超螺旋程度的拓樸異構酶;還有多種調節基因表現的轉錄因子。
前體mRNA的處理
新合成的mRNA分子稱為初級轉錄產物或前體mRNA,在送入細胞質以前,必須先在細胞核內經過後轉錄修飾作用的處理。而細胞核內未受修飾的mRNA,將會被降解而非用作蛋白質的轉譯合成。三種主要的修飾作用是5'端加帽、3'端的多聚腺苷酸化,以及RNA剪接作用。前體mRNA會在細胞核中與多種蛋白質複合成異源核糖核蛋白顆粒(hnRNP)。加上5'端帽的過程與轉錄作用同時進行,是後轉錄修飾的第一個步驟。3'端多聚腺苷酸尾鏈則是在轉錄完成後才會加上。
RNA剪接是由稱為剪接體的複合物進行,此過程中前體mRNA上不會轉譯成蛋白質的內含子將會被移除,之後將切割過的外顯子片段重新組合成連續的分子。剪接作用通常發生在5'端帽與3'多聚腺苷酸處理完成之後,不過在一些外顯子也可以發生在轉錄完成以前。包括可編碼出抗體的許多前體mRNA,可以經由多種不同的剪接方式生產出不同的mRNA,進而轉錄出多種不同的蛋白質序列。這種過程稱為選擇性剪接,使一段變化有限的DNA得以生產出許多不一樣的蛋白質。
無核與多核細胞
雖然多數細胞都有一個細胞核,但也有些細胞沒有細胞核,還有一些則是擁有多個細胞核。這可能屬於正常現象,如哺乳類的紅血球;也可能是肇因於細胞分裂過程中的不正常錯誤。另外原核细胞雖然没有细胞核,但有染色较深,含DNA多的区域,称为拟核。
無核細胞沒有細胞核,因此不具有分裂並製造姊妹細胞的能力。了解最透徹的無核細胞是哺乳類的紅血球,這種細胞也少了其他的胞器,如線粒體。紅血球主要的功能是作為運輸工具,將肺部裡的氧氣送往身體各處組織。紅血球是在骨髓中經由紅血球生成作用產生,並在此過程中失去細胞核、胞器,以及核糖體。細胞核是在紅血球母細胞分化形成網狀紅血球,也就是形成成熟紅血球前體的過程中遭到排除。當存在某些突變原時,則可能導致部份未成熟的「微核」紅血球被釋放到血流當中。除此之外,無核細胞也可能在錯誤的細胞分裂中產生,此時兩個姊妹細胞中有一個無核,另一個則有兩個核。
多核細胞含有多個細胞核。原生動物中多數屬於等輻骨蟲的物種,以及部分真菌類的菌根裡,有自然形成的多核細胞。而人類骨骼肌中的肌細胞,也會在發育過程中形成多核細胞。這些細胞核排列在在靠近細胞邊緣的位置,產生最大的細胞內空間供肌原纖維通過。人體中有一些不正常形成的多核細胞,例如當單核球與巨噬細胞融合時,會產生巨型多核細胞,有時會伴隨著發炎反應,並與腫瘤的形成有關。
演化
細胞核是真核細胞的主要結構,也因此有許多關於演化起源的推測。有四種主要理論可解釋細胞核的存在,而這些理論皆尚未受到廣泛支持。
「共營模型」(syntrophic model)認為,古菌與細菌的共生,導致了含細胞核的真核細胞誕生。類似於現代產甲烷古菌的某些古代古菌,侵入並生活在類似於現代粘細菌的細菌體內,形成了早期的細胞核。此理論類似於原始真核生物與好氧細菌的內共生關係,也就是解釋粒線體與葉綠體起源的理論。古菌與真核生物在特定蛋白質,如組織蛋白基因的相似性,支持了以古菌為基礎的細胞核起源理論。觀察顯示黏細菌可自行運動,並形成多細胞複合體,也擁有與真核生物相似的激酶與G蛋白,此現象支持了真核細胞起源於細菌的說法。
第二種模型認為,原始的真核細胞是在未發生內共生的狀況下,自細菌演化而來。此理論的基礎在於現代浮黴細菌擁有包含原始核孔與其他分隔模構造的核狀結構。另一項類似說法指出,一種稱為慢性細胞(chronocyte)的類真核細胞,首先演化成型,並將古菌與細菌吞噬到體內,使細胞核與真核細胞形成。
還有一項較具爭議性的模型,稱為「病毒性真核生物起源」(viral eukaryogenesis),此模型認為病毒感染了原核生物,導致膜結合細胞核與其他真核生物特徵的成型。這種理論的基礎在於真核生物與病毒間的某些相似性,如線性DNA、mRNA的加帽作用,以及蛋白質的緊密結合(病毒的外套膜相當於真核生物的組織蛋白)。理論的其中一個版本認為,吞噬作用形成了早期的細胞「掠食者」,並因此演化出細胞核。也有理論認為真核生物起源於受到痘病毒感染的古菌;因為觀察顯示,現代痘病毒與真核生物的DNA聚合酶具相似性。此外,病毒性真核生物起源假說,也可與部分性別相關演化問題有關。
一項近期研究顯示,傳統上類似於內共生學說的觀點,不足以有效地解釋真核生物細胞核的起源。新模型稱作「外膜假說」(exomembrane hypothesis),認為細胞核是起源自演化出第二層外細胞膜的早期細胞;其靠近內部的膜轉變成為細胞核膜,並逐漸演化出複雜的核孔結構,以幫助如核糖體單元等內部核成的物質送出核外。