葉綠體
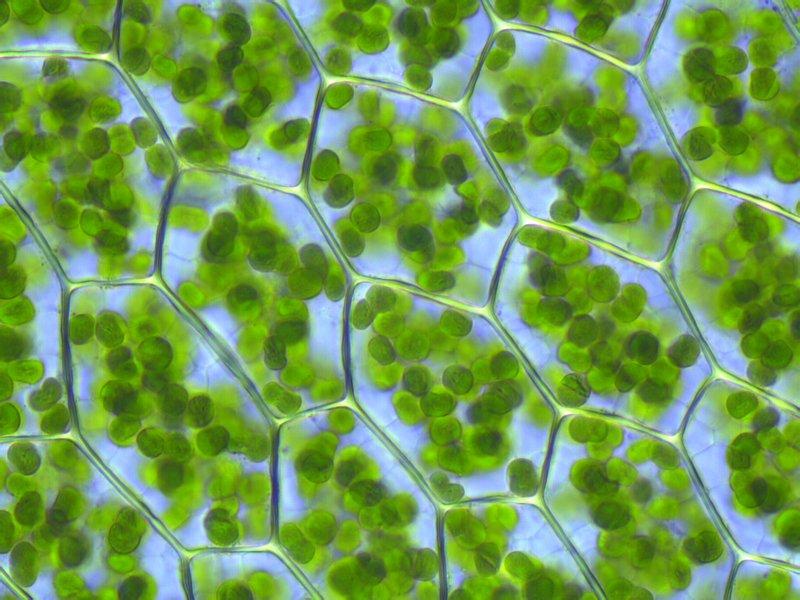
葉綠體 ( chloroplast )[1]是植物細胞內最重要、最普遍的質體,它是進行光合作用的細胞器。是綠色植物和藻類等真核自養生物細胞中專業化亞單元的細胞器。葉綠體利用其葉綠素將光能轉變為化學能,把CO2與水轉變為糖。葉綠體是世界上成本最低、創造物質財富最多的生物工廠。
目錄
基本介紹
- 中文學名:葉綠體
- 拉丁學名:chloroplast
- 界:植物界
- 分布區域:植物莖葉
- 長徑:視情況而定 5~100μm不等
- 主要:葉綠素和細胞素
- 主要作用:進行光合作用
葉綠體介紹
大部分高等植物和藻類微生物的葉綠體內類囊體緊密堆積。主要含有葉綠素(葉綠素a和葉綠素b)、類胡蘿蔔素(胡蘿蔔素和葉黃素),葉綠素a和葉綠素b主要吸收藍紫光和紅光,胡蘿蔔素和葉黃素主要吸收藍紫光。這些色素吸收的光都可用於光合作用,葉綠素在色素所占比例最大,且吸收綠光最少,因此綠光被反射,細胞呈現綠色。葉綠體(chloroplast)存在於藻類和綠色植物中的色素體之一,光合作用的生化過程在其中進行。因為葉綠體除含黃色的胡蘿蔔素外,還含有大量的葉綠素,所以看上去是綠色的。褐藻和紅藻的葉綠體除含葉綠素外還含有藻黃素和藻紅蛋白,看上去是褐色或紅色[有人分別稱為褐色體(phacaplost)、紅色體 (rhodoplast)]。許多植物的葉綠體是直徑5微米左右,厚2—3微米的凸透鏡形狀,但低等植物中則含有板狀、網眼狀、螺旋形、星形、杯形等非常大的葉綠體。葉肉細胞中含的葉綠體數通常是數十到數百個。已知有的一個細胞含有數千個以上葉綠體的例子,以及僅有一個葉綠體的例子。用光學顯微鏡觀察葉綠體,它的平面相多數為0.5微米大小的濃綠色粒狀結構(基粒)。基粒的清晰程度和數量隨植物和組織的種類及葉綠體的發育時期而不同,反映著內膜系統的分化程度。包著葉綠體的包膜由內外兩層膜組成,對各種各樣的離子以及種種物質具有選擇透過性。在葉綠體內部有基質、富含脂質和質體醌的質體顆粒,以及結構精細的內膜系統(片層構造,類囊體)。在基質中水占葉綠體重量的60%—80%,這裡有各種各樣的離子、低分子有機化合物、酶、蛋白質、核糖體、RNA、DNA等。在綠藻、褐藻,紅藻、接合藻、硅藻等許多藻類的葉綠體中存在著澱粉核。構成內膜系統微細結構基礎的是類囊體。在具有基粒的葉綠體中重疊起類囊體或複雜地摺疊起來,分化成所謂的基粒堆(grana stack)和與之相聯繫的膜系統[基粒間片層(intergrana lamellae)]。各種光合色素和光合成電子傳遞成分、磷酸化偶聯因子等存在於類囊體中,色素被光能激發、電子傳遞、直到ATP合成都在類囊體上及其表面附近進行。利用由此生成的NADPH和ATP在基質中進行二氧化碳固定。
幾乎可以說一切生命活動所需的能量來源於太陽能(光能)。綠色植物是主要的能量轉換者是因為它們均含有葉綠體(Chloroplast)這一完成能量轉換的細胞器,它能利用光能同化二氧化碳和水,合成貯藏能量的有機物,同時產生氧。所以綠色植物的光合作用是地球上有機體生存、繁殖和發展的根本源泉。
- 葉綠體

葉綠體(chloroplast)和粒線體類似,[2]都是由向內摺疊的雙層膜所構成,但葉綠體是植物細胞特有的構造,內部含有葉綠素,可進行光合作用,用二氧化碳和水分子為原料,經過一連串的化學反應,將太陽光能轉變為儲存在葡萄糖分子內的化學能,同時釋放出氧氣。藉由光合作用所合成的葡萄糖分子,在植物細胞內可進一步聚合反應,形成澱粉和纖維素等各種多醣類。
在高解析度的光學顯微鏡下,高等植物的葉綠體呈圓盤形,而根據解析度更高的電子顯微鏡,可觀察到葉綠體是一種複雜的板狀(lamella)構造,一層層排列緊密的板狀部分稱為葉綠餅(granum),而圍繞著葉綠餅的物質則稱作基質(stroma),葉綠餅與葉綠餅之間有許多貫串其間的膜板連接,葉綠素捕捉光能的機制,便與葉綠素在葉綠餅板層上的分布情形有關。由於葉綠體可將光能轉換為生物體可利用的能量,因此也被稱為「細胞的太陽能工廠」。葉綠體可能起源於古代藍藻,因為藍藻中有葉綠素。某些古代真核生物靠吞噬其他生物維生,它們吞下的某些藍藻沒有被消化,反而依靠吞噬者的生活廢物製造營養物質。
高等植物的葉綠體存在於細胞質基質中。葉綠體一般是綠色的扁平的快速流動的橢球形或球形,可以用高倍光學顯微鏡觀察它的形態和分布。
形態與結構
在高等植物中葉綠體象雙凸或平凸透鏡,長徑5~10um,短徑2~4um,厚2~3um。高等植物的葉肉細胞一般含50~200個葉綠體,可占細胞質的40%,葉綠體的數目因物種細胞類型,生態環境,生理狀態而有所不同。在藻類中葉綠體形狀多樣,有網狀、帶狀、裂片狀和星形等等,而且體積巨大,可達100um。
葉綠體由葉綠體外被(chloroplast envelope)、類囊體(thylakoid)和基質(stroma)3部分組成,葉綠體含有3種不同的膜:外膜、內膜、類囊體膜和3種彼此分開的腔:膜間隙、基質和類囊體腔。
- 外被
葉綠體外被由雙層膜組成,膜間為10~20nm的膜間隙。外膜的滲透性大,如核苷、無機磷、蔗糖等許多細胞質中的營養分子可自由進入膜間隙。
內膜對通過物質的選擇性很強,CO2、O2、Pi、H2O、磷酸甘油酸、丙糖磷酸,雙羧酸和雙羧酸胺基酸可以透過內膜,ADP、ATP已糖磷酸,葡萄糖及果糖等透過內膜較慢。蔗糖、C5糖雙磷酸酯,C糖磷酸酯,NADP+及焦磷酸不能透過內膜,需要特殊的轉運體(translator)才能通過內膜。
- 類囊體
是單層膜圍成的扁平小囊,沿葉綠體的長軸平行排列。膜上含有光合色素和電子傳遞鏈組分,又稱光合膜。
許多類囊體象圓盤一樣疊在一起,稱為基粒,組成基粒的類囊體,叫做基粒類囊體,構成內膜系統的基粒片層(grana lamella)。基粒直徑約0.25~0.8μm,由10~100個類囊體組成。每個葉綠體中約有40~60個基粒。
葉綠體通過內膜形成類囊體來增大內膜面積,以此為在葉綠體中發生的反應提供場所。
貫穿在兩個或兩個以上基粒之間的沒有發生垛疊的類囊體稱為基質類囊體,它們形成了內膜系統的基質片層(stroma lamella)。
由於相鄰基粒經網管狀或扁平狀基質類囊體相聯結,全部類囊體實質上是一個相互貫通的封閉系統。類囊體做為單獨一個封閉膜囊的原始概念已失去原來的意義,它所表示的僅僅是葉綠體切面的平面形態。
類囊體膜的主要成分是蛋白質和脂類(60:40),脂類中的脂肪酸主要是不飽和脂肪酸(約87%),具有較高的流動性。光能向化學能的轉化是在類囊體上進行的,因此類囊體膜亦稱光合膜,類囊體膜的內在蛋白主要有細胞色素b6/f複合體、質體醌(PQ)、質體藍素(PC)、鐵氧化還原蛋白、黃素蛋白、光系統Ⅰ、光系統Ⅱ複合物等。
- 基質
是內膜與類囊體之間的空間的液體,主要成分包括:
碳同化相關的酶類:如RuBP羧化酶占基質可溶性蛋白總量的60%。
葉綠體DNA、蛋白質合成體系:如,ctDNA、各類RNA、核糖體等。
一些顆粒成分:如澱粉粒、質體小球和植物鐵蛋白等。葉綠體的功能葉綠體(chloroplast):藻類和植物體中含有葉綠素進行光合作用的器官。 主要含有葉綠素、胡蘿蔔素和葉黃素,其中葉綠素的含量最多,遮蔽了其他色素,所以呈現綠色。主要功能是進行光合作用。幾乎可以說一切生命活動所需的能量來源於太陽能(光能)。綠色植物是要的能量轉換者是因為它們均含有葉綠體(Chloroplast)這一完成能量轉換的細胞器。
- 光合作用
光合作用的是能量及物質的轉化過程。首先光能轉化成電能,經電子傳遞產生ATP和NADPH形式的不穩定化學能,最終轉化成穩定的化學能儲存在糖類化合物中。分為光反應(light dependent reaction)和暗反應(light independent reaction),前者需要光,涉及水的光解和光合磷酸化,後者不需要光,涉及CO2的固定。分為C3和C5兩類。暗反應需要光反應產生的能量來進行。
1、光合色素
類囊體中含兩類色素:葉綠素和橙黃色的類胡蘿蔔素,通常葉綠素和類胡蘿蔔素的比例約為3:1,chla與chlb葉綠素的實驗中,隨層析液在濾紙上擴散最快的是胡蘿蔔素
2、集光複合體(light harve為3:l,全部葉綠素和幾乎所有的類胡蘿蔔素都包埋在類囊體膜中,與蛋白質以非共價鍵結合,一條肽鏈上可以結合若干色素分子,各色素分子間的距離和取向固定,有利於能量傳遞。
在提取和分離葉綠體中色素的實驗中,隨層析液在濾紙上擴散最快的是胡蘿蔔素
3、集光複合體(light harvesting complex)
由大約200個葉綠素分子和一些肽鏈構成。大部分色素分子起捕獲光能的作用,並將光能以誘導共振方式傳遞到反應中心色素。因此這些色素被稱為天線色素。葉綠體中全部葉綠素b和大部分葉綠素a都是天線色素。另外類胡蘿蔔素和葉黃素分子也起捕獲光能的作用,叫做輔助色素。
4、細胞色素b6/f複合體(cyt b6/f complex) 可能以二聚體形成存在,每個單體含有四個不同的亞基。細胞色素b6(b563)、細胞色素f、鐵硫蛋白、以及亞基Ⅳ(被認為是質體醌的結合蛋白)。
5、光系統Ⅰ(PSI)
能被波長700nm的光激發,又稱P700。包含多條肽鏈,位於基粒與基質接觸區和基質類囊體膜中。由集光複合體Ⅰ和作用中心構成。結合100個左右葉綠素分子、除了幾個特殊的葉綠素為中心色素外,其它葉綠素都是天線色素。三種電子載體分別為A0(一個chla分子)、A1(為維生素K1)及3個不同的4Fe-4S。
6、光系統Ⅱ(PSⅡ)
吸收高峰為波長680nm處,又稱P680。至少包括12條多肽鏈。位於基粒與基質非接觸區域的類囊體膜上。包括一個集光複合體(light-hawesting comnplex Ⅱ,LHC Ⅱ)、一個反應中心和一個含錳原子的放氧的複合體(oxygen evolving complex)。D1和D2為兩條核心肽鏈,結合中心色素P680、去鎂葉綠素(pheophytin)及質體醌(plastoquinone)。
- 傳遞方式
P680接受能量後,由基態變為激發態(P680*),然後將電子傳遞給去鎂葉綠素(原初電子受體),P680*帶正電荷,從原初電子供體Z(反應中心D1蛋白上的一個酪氨酸側鏈)得到電子而還原;Z+再從放氧複合體上獲取電子;氧化態的放氧複合體從水中獲取電子,使水光解。
2H2O→O2+4H++4e-
在另一個方向上去鎂葉綠素將電子傳給D2上結合的QA,QA又迅速將電子傳給D1上的QB,還原型的質體醌從光系統Ⅱ複合體上游離下來,另一個氧化態的質體醌占據其位置形成新的QB。質體醌將電子傳給細胞色素b6/f複合體,同時將質子由基質轉移到類囊體腔。電子接著傳遞給位於類囊體腔一側的含銅蛋白質體藍素(plastocyanin, PC)中的Cu2+,再將電子傳遞到光系統Ⅱ。
P700被光能激發後釋放出來的高能電子沿著A0→ A1 →4Fe-4S的方向依次傳遞,由類囊體腔一側傳向類囊體基質一側的鐵氧還蛋白(ferredoxin,FD)。最後在鐵氧還蛋白-NADP還原酶的作用下,將電子傳給NADP+,形成NADPH。失去電子的P700從PC處獲取電子而還原
以上電子呈Z形傳遞的過程稱為非循環式光合磷酸化,當植物在缺乏NADP+時,電子在光系統內Ⅰ流動,只合成ATP,不產生NADPH,稱為循環式光合磷酸化。
- 光合磷酸化
一對電子從P680經P700傳至NADP+,在類囊體腔中增加4個H+,2個來源於H2O光解,2個由PQ從基質轉移而來,在基質外一個H+又被用於還原NADP+,所以類囊體腔內有較高的H+(pH≈5,基質pH≈8),形成質子動力勢,H+經ATP成合酶,滲入基質、推動ADP和Pi結合形成ATP。
ATP合成酶,即CF1-F0偶聯因子,結構類似於線粒體ATP合成酶。CF1同樣由5種亞基組成α3β3γδε的結構。CF0嵌在膜中,由4種亞基構成,是質子通過類囊體膜的通道。
- 碳反應
國際通用名稱為碳反應,而非暗反應。因為該反應在沒有光的時候,會因為缺乏光反應產生的ATP及NADPH而無法進行。
C3途徑(C3 pathway):亦稱卡爾文(Calvin)循環。CO2受體為RuBP,最初產物為3-磷酸甘油酸(PGA)。
C4途徑(C4 pathway):亦稱哈奇-斯萊克(Hatch-Slack)途徑,CO2受體為PEP,最初產物為草醯乙酸(OAA)。
景天科酸代謝途徑(Crassulacean acid metabolism pathway,CAM途徑):夜間固定CO2產生有機酸,白天有機酸脫羧釋放CO2,進行CO2固定。
- 半自主性
線粒體與葉綠體都是細胞內進行能量轉換的場所,兩者在結構上具有一定的相似性。①均由兩層膜包被而成,且內外膜的性質、結構有顯著的差異。②均為半自主性細胞器,具有自身的DNA和蛋白質合成體系。因此綠色植物的細胞記憶體在3個遺傳系統。
葉綠體DNA由Ris和Plaut 1962最早發現於衣藻葉綠體。ctDNA呈環狀,長40~60μm,基因組的大小因植物而異,一般約200bp-2500bp。數目的多少植物的發育階段有關,如菠菜幼苗葉肉細胞中,每個細胞含有20個葉綠體,每個葉綠體含DNA分子200個,但到接近成熟的葉肉細胞中有葉綠體150個,每個葉綠體含30個DNA分子。
和線粒體一樣,葉綠體只能合成自身需要的部分蛋白質,其餘的是在細胞質游離的核糖體上合成的,必需運送到葉綠體,才能發揮葉綠體應有的功能。已知由ctDNA編碼的RNA和多肽有:葉綠體核糖體中4種rRNA(23S、16S、4.5S及5S),20種(菸草)或31種(地錢)tRNA,約90多種多肽。
由於葉綠體在形態、結構、化學組成、遺傳體系等方面與藍細菌相似,人們推測葉綠體可能也起源於內共生的方式,是寄生在細胞內的藍藻演化而來的。
與質體區別
葉綠體含有四種色素:葉綠素a、葉綠素b、葉黃素及胡蘿蔔素。其中前二者為主要的光合色素,直接參與光合作用;後二者僅起吸收、傳遞光能的作用,而不能參與光合作用。由於不同植物體中或同一植物不同發育時期,細胞中所含四種色素的比例不斷變化,因而植物(尤其為葉)在顏色上表現出深淺黃綠色的不同。
葉綠體的內部結構十分複雜。在電鏡下觀察葉綠體,可見,葉綠體的外表是由雙層平滑的單位膜構成的葉綠體被膜(chloroplastenvelop),其內是無色的基質(matrix)(其主要成分是親水的蛋白質),基質中分布著若干個含有葉綠素的基粒(granum)。這些基粒是由許多層迭合的片層結構組成,此片層結構稱為類囊體(thylokoid)。類囊體由單層膜圍合而成,其上分布有許多穿孔。囊內含有液狀的內含物。類囊體除平行垛迭構成基粒外,還在基質內到處延伸,從而構成了複雜的類束體系統(thylokoidsystem)。其中構成基粒的類囊體部分稱基粒片層(granalamella),而連線基粒的類囊體部分,稱為基質片層(stromalamella)。
葉綠體所含的色素存在於類囊體膜上,與蛋白質結合形成複合體(complax),包藏或連線在類囊體膜的磷脂分子的雙層中,光合作用就在這裡進行。
在個體發育中,葉綠體來自前質體(proplastid)—即未分化的質體,存在於根尖、莖尖,其結構較為簡單。
在直接光照下,幼葉中的前質體,內層膜在許多部位內折而伸入基質中,並逐漸擴展增大,最終脫離內層膜,形成扁平的囊狀結構——類囊體。許多個類束體垛合在一起便形成為基粒,由此前質體逐步地發育成為成熟的葉綠體。但在黑暗或光照不足的情況下,就不能形成正常的類囊體系統,而形成由許多小泡組成的格線狀結構,稱前片層體(prollamellabody)。這樣的質體稱為黃化體(etioplast)。一般在獲取光照後,黃化體中的前片層體可進一步轉變,發育成為具有基粒結構的正常葉綠體。
有色體(chromoplast):主要存在於花瓣、果實、貯藏根及衰老的葉片中,主要功能尚不十分清楚,但有一點是明確的,即積聚澱粉和脂類,並可幫助傳粉。
有色體所含色素主要是葉黃素和胡蘿蔔素,並因所含色素比例的不同而呈現紅色——黃色之間的色彩梯度變化。
有色體即可由前質體發育而來,也可由葉綠體失去葉綠素轉化而來,如果實的成熟即為一實例,另外,還可由白色體轉化而來,胡蘿蔔即為如此。
有色體的形狀是多種多樣的。
白色體(leucoplast):是不含可見色素的無色質體,呈顆粒狀。存在於一些植物的貯藏器官中,如甘薯、土豆的地下器官及種子的胚中。
白色體的主要功能是積累澱粉、蛋白質及脂肪,從而使其相應地轉化為澱粉粒、糊粉粒和油滴。
應注意的是,白色體雖不含可見色素,卻含無色的原葉綠素,故見光後便可轉化為葉綠體。如土豆風吹後變綠的現象即為如此。
以上兩種質體,即有色體和白色體的結構雖不及葉綠體,但也較複雜。表面均為雙層單位膜構成的質體被膜,內有以親水蛋白質為主的液狀基質,由於沒有基粒結構存在,無發達的類囊體系統而區別於葉綠體。
總之,質體是一類合成和積累同化產物的細胞器。
- 小提示:
質體體積較小,呈園盤形(扁園形)或扁卵園形,直徑約為5-8um,厚約1um。
根據顏色及功能的不同,質體可分為三種:葉綠體、白色體及有色體。
增殖
在個體發育中葉綠體由原質體發育而來,原質體存在於根和芽的分生組織中,由雙層被膜包圍,含有DNA,一些小泡和澱粉顆粒的結構,但不含片層結構,小泡是由質體雙層膜的內膜內折形成的。
在有光條件原質體的小泡數目增加並相互融合形成片層,多個片層平行排列成行,在某些區域增殖,形成基粒,變成綠色原質體發育成葉綠體。
在黑暗性長時,原質體小泡融合速度減慢,並轉變為排列成格線的小管的三維晶格結構,稱為原片層,這種質體稱為黃色體。黃色體在有光的情況下原片層彌散形成類囊體,進一步發育出基粒,變為葉綠體。
葉綠體能靠分裂而增殖,這各分裂是靠中部縊縮而實現的,在發育7天的幼葉的基部2-2.5cm處很容易看到幼齡葉綠體呈啞鈴形狀,從菠菜幼葉含葉綠體少、ctDNA多,老葉含葉綠體多,每個葉綠體含ctDNA少的現象也可以看出葉綠體是以分裂的方式增殖的。
成熟葉綠體正常情況下一般不再分裂或很少分裂。
高等植物的葉綠體主要存在於葉肉細胞內,含有葉綠素。電鏡觀察表明:葉綠體外有光滑的雙層單位膜,內膜向內疊成類囊體,若干類囊體垛疊成基粒。基粒內的某些類囊體內向外伸展,連線不同基粒。連線基粒的類囊體部分,稱為基質片層;構成基粒的類囊體部分,稱為基粒片層。
在個體發育上,葉綠體來自前質體,由前質體發育成葉綠體。並且,無光不能形成葉綠素。
- 發現
1880年,法國植物學家、植物葉綠體的發現者席姆佩爾證明澱粉是植物光合作用的產物。1883年,他經研究發現澱粉只在植物細胞的特定部位形成,並將其命名為葉綠體,席姆佩爾還曾廣泛地遊歷了美洲、亞洲和非洲的熱帶地區,對那裡的植物進行了考察,並將考察結果發表在1898年出版的《以生理學為基礎的植物--------地理學》一書中。
1940年,德國人G.A.Kausche和H.Ruska發表了世界第一張葉綠體的電鏡照片。
- 起源
(一)內共生起源學說 許多科學家認為,線粒體和葉綠體分別起源於原始真核細胞內共生的細菌和藍藻。1970年Margulis在分析了大量資料的基礎上提出了一種構想,認為真核細胞的祖先是一種體積巨大的、不需氧的、具有吞噬能力的細胞,能將吞噬所得的糖類進行酵解取得能量。而線粒體的祖先——原線粒體則是一種革蘭氏陰性菌,含有三羧酸循環所需的酶系和電子傳遞鏈,故它可利用氧氣把糖酵解的產物丙酮酸進一步分解,獲得比酵解更多的能量。當這種細菌被原始真核細胞吞噬後,即與宿主細胞間形成互利的共生關係,原始真核細胞利用這種細菌(原線粒體)充分供給能量,而原線粒體從宿主細胞獲得更多的原料。
(二)非共生起源學說 該學說的支持者提出一種線粒體和葉綠體起源的構想,認為真核細胞的前身是一個進化上比較高等的好氧細菌,它比典型的原核細胞大,這樣就要逐漸增加具有呼吸功能的膜表面,開始是通過細菌的細胞膜內陷、擴張和分化,後逐漸形成了線粒體和葉綠體的雛形。根據1974年Uzzell等人的觀點,在進化的最初階段,原核細胞的基因組進行複製並不伴有細胞分裂,然後基因附近的質膜內陷形成雙層膜,分別將基因組包圍在這些雙層膜結構中,從而形成了原始線粒體、葉綠體等細胞器。後來在進化過程中進一步發生了分化,如線粒體和葉綠體的基因組丟失一些基因;細胞核的基因則有了高度發展;質體發展了光合作用;線粒體則演變為專具有呼吸功能的細胞器,於是逐漸形成了現在的真核細胞。
從目前看,對這兩個學說尚有爭議,各有其實驗證據和支持者,因此,關於線粒體和葉綠體的起源,有待今後進一步探討和研究。
運動

在暗環境或者弱光的照射下,葉綠體會較分散的分布在細胞各處。[3]但在強光的照射下,為了更好地利用光能,葉綠體會游移依附到細胞膜下,一方面減少相互之間的重疊,另一方面可以因減少了光經過的路程而得到更大的光強度。