細胞骨架
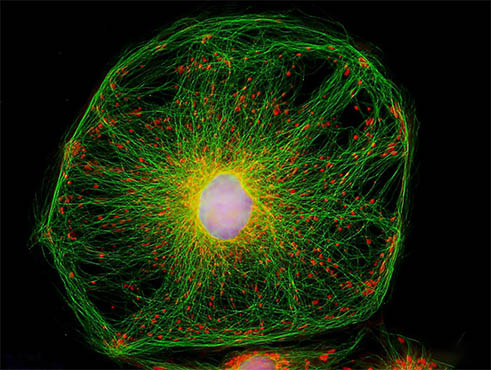 |
細胞骨架是真核細胞中的蛋白纖維網架體系。
狹義的細胞骨架(cytoskeleton)概念是指真核細胞中的蛋白纖維網架體系( 微管(microtubule, MT)、微絲(microfilament, MF)及中間纖維(intermediate filament, IF )組成的體系),[1]它所組成的結構體系稱為「細胞骨架系統」,與細胞內的遺傳系統、生物膜系統、並稱「細胞內的三大系統」。直到20世紀60年代後,採用戊二醛常溫固定,才逐漸認識到細胞骨架的客觀存在,它是真核細胞藉以維持其基本形態的重要結構,被形象地稱為細胞骨架,它通常也被認為是廣義上細胞器的一種。廣義的細胞骨架概念是細胞核骨架、細胞質骨架、細胞膜骨架和胞外基質所形成的網絡體系。核骨架、核纖層與中間纖維在結構上相互連接,貫穿於細胞核和細胞質的網架體系。[2]
目錄
作用
細胞骨架(cytoskeleton)是指真核細胞中的蛋白纖維網絡結構,發現較晚,主要是因為一般電鏡制樣採用低溫(0-4℃)固定,而細胞骨架會在低溫下解聚。
直到20世紀60年代後,採用戊二醛常溫固定,才逐漸認識到細胞骨架的客觀存在。真核細胞藉以維持其基本形態的重要結構,被形象地稱為細胞骨架,它通常也被認為是廣義上細胞器的一種。
細胞骨架不僅在維持細胞形態,承受外力、保持細胞內部結構的有序性方面起重要作用,而且還參與許多重要的生命活動,如:在細胞分裂中細胞骨架牽引染色體分離,在細胞物質運輸中,各類小泡和細胞器可沿着細胞骨架定向轉運;在肌肉細胞中,細胞骨架和它的結合蛋白組成動力系統;在白細胞(白血球)的遷移、精子的遊動、神經細胞軸突和樹突的伸展等方面都與細胞骨架有關。另外,在植物細胞中細胞骨架指導細胞壁的合成。
微管
微管(microtubule)可在所有哺乳類動物細胞中存在,直徑大於12nm,除了紅細胞(紅血球)外,所有微管均由約55kD的α及β微管蛋白(tubulin)組成。它們正常時以(αβ)二聚體形式存在,並以頭尾相連的方式聚合,形成微管蛋白原纖維(protofilament),一般由13根這樣的原纖維構成一個中空的微管,直徑22~25nm。少數變異的微管如線蟲等所有的則有其他數目的原纖維。微管確定膜性細胞器(membrane-enclosed organelle)的位置和作為膜泡運輸的導軌。微管是細胞骨架的架構主幹,並也是某些胞器的主體,例如中心粒(centriole)就是由9組3聯微管組成的構造,而真核生物的纖毛(cilium)與鞭毛(flagellum)也是由以微管為9+2結構,即由9個二聯微管和一對中央微管構成,其中二聯微管由AB兩個管組成,A管由13條原纖維組成,B管由10條原纖維組成,兩者共享5條。A管對着相鄰的B管伸出兩條動力蛋白臂,並向鞭毛中央發出一條輻。基體的微管組成為9+0,並且二聯微管為三聯微管所取代。組成的軸絲(axoneme)為主體。
從各種組織中提純微管蛋白可以發現還存在一些其他蛋白成分(5%-20%),稱之謂微管相關蛋白(microtube associated proteins MAPs)。這些蛋白具有組織特異性,表現出從相同αβ二聚體聚合形成的微管具有獨特的性質,已從人類不同組織中發現了多種α及β微管蛋白,並追蹤微管基因表現出部分基因家族,某些基因被認為是編碼獨特的微管蛋白。
微管形成的有些結構是比較穩定的,是由於微管結合蛋白的作用和酶修飾的原因。如神經細胞軸突、纖毛和鞭毛中的微管纖維。大多數微管纖維處於動態的聚合和災變(一種突然的,迅速的,一般不可逆轉的分解)狀態,這是實現其功能所必需的性質(如紡錘體)。與秋水仙素(colchicine)結合的微管蛋白可加合到微管上,並阻止其他微管蛋白單體繼續添加,進而破壞紡錘體的結構,長春花鹼具有類似的功能。紫杉酚(taxol),能促進微管的聚合,並使已形成的微管穩定,然而這種穩定性會破壞微管的正常功能。這些藥物可以利用破壞微管功能以阻止細胞分裂,成為癌症治療的新希望。
在人類至少發現兩種明顯區別的α-微管蛋白及三種明顯區別的β-微管基因,它們產生具有特定功能的微管蛋白mRNA,由於這些編碼在結構組分上十分近似蛋白質分子,在不同組織存在多少特異性的具有差異表達的微管蛋白亞型,尚待深入研究。
除了α-與β-微管蛋白有編碼相似的不同變異型,近幾年來又發現了多種編碼差異更大的新的微管蛋白,形成不同的基因家族。其中gamma微管蛋白位於細胞內的微管組織中心(microtubule organizing center, MTOC),是用以提供α及β微管蛋白進行聚合反應形成微管的起始核心。而delta與epsilon則被認為與中心體(centrosome)的結構與形成有關。其他尚有eta, zeta, theta等等多種變異,不過通常僅存在少數幾種真核單細胞生物如原蟲或纖毛蟲里,可能跟這些生物獨特的結構與生理習性有關,進一步詳情仍待研究 。
微絲
微絲(microfilament)也普遍存在於所有真核細胞中,是一個實心狀的纖維,直徑為4nm-7nm一般細胞中含量約占細胞內總蛋白質的1%-2%,但在活動較強的細胞中可占20%-30%。在一般細胞主要分布於細胞的表面,直接影響細胞的形狀。微絲具有多種功能,在不同細胞的表現不同,在肌細胞組成粗肌絲、細肌絲,可以收縮(收縮蛋白),在非肌細胞中主要起支撐作用、非肌性運動和信息傳導作用。
微絲主要由肌動蛋白(actin)構成,和肌球蛋白(myosin,一種分子馬達蛋白)一起作用,使細胞運動。它們參與細胞的變形蟲運動、植物細胞的細胞質流動與肌肉細胞的收縮:
植物細胞的細胞質流動: 微絲中的actin(肌動蛋白)與myosin(肌球蛋白)在細胞質形成三維的網絡體系。actin位於外質,myosin位於內質。 myosin連結着細胞質顆粒,由ATP供給能量,myosin與細胞質顆粒的結合體沿着actin filament滑動,從而帶動整個細胞質的環流。
變形蟲運動(amoeboid movememt,阿米巴運動): 肌肉細胞的收縮:
如同微管蛋白,肌動蛋白的基因組成一個超家族,並組成多種極為相似的結構。例如,各種肌肉細胞有不同的肌動蛋白:①骨骼肌的條紋纖維;②心肌的條紋纖維;③血管壁的平滑肌;④胃腸道壁的平滑肌。它們在氨基酸組分上有微小的差異(大約在400個氨基酸殘基序列中有4-6個變異),在肌肉與非肌細胞中都還存在β及γ肌動蛋白,它們與具有橫紋的α肌動蛋白可有25個氨基酸的差異。
G-肌動蛋白單體(含ATP)可聚合為呈纖維狀的F-肌動蛋白(含ADP),它們可由Mg2+及高濃度的K+或Na+誘導而聚合,聚合後ATP水解為ADP及C-肌動蛋白ADP單體,組成F-肌動蛋白。在骨骼肌的細肌絲(thin filament,由肌動蛋白構成)與粗肌絲(thick filament,由肌球蛋白構成)相互作用而使肌肉收縮(肌球蛋白可以起作肌動蛋白激活的ATPase的作用)。肌球蛋白也存在於哺乳動物的非肌細胞中(但以非聚合狀態存在)。
中間纖維
細胞骨架的第三種纖維結構稱中間纖維(intermediate filament,IF),又稱中間絲、中等纖維,直徑介於微管和微絲之間(8nm-10nm),其化學組成比較複雜。構成它的蛋白質多達5種,常見的有波形蛋白(vimentin)、角蛋白(keratin)、結蛋白、神經元纖維、神經膠質纖維。在不同細胞中,成分變化較大。中間纖維使細胞具有張力和抗剪切力。中間纖維有共同的基本結構,即構建成一個中央α螺旋杆狀區,兩側則是大小和化學組成不同的端區。端區的多樣性決定了中間纖維外形和性質的差異和特異性。
以上這些結構單元並非是一成不變的,而是隨細胞的生命活動而呈現高度的動態性,它們均由單體蛋白以較弱的非共價鍵結合在一起,構成纖維型多聚體,很容易進行組裝和去組裝,這正是實現其功能所必需的特點。
原核生物
長期以來,人們認為細胞骨架僅為真核生物所特有的結構,但近年來的研究發現它也存在於細菌等原核生物中。
人們已經在細菌中發現的FtsZ、MreB 和CreS 依次與真核細胞骨架蛋白中的微管蛋白、肌動蛋白絲及中間絲類似。FtsZ 能在細胞分裂位點裝配形成Z 環結構,並通過該結構參與細胞分裂的調控;MreB能形成螺旋絲狀結構,其主要功能有維持細胞形態、調控染色體分離等;CreS存在於新月柄桿菌中,它在細胞凹面的細胞膜下面形成彎曲絲狀或螺旋絲狀結構,該結構對維持新月柄桿菌細胞的形態具有重要作用。
疾病及危害
細胞骨架是細胞生命活動中不可缺少的細胞結構,其形成的複雜網絡體系對細胞形態的改變和維持、細胞的分裂與分化、細胞內物質運輸、細胞信息傳遞、基因表達等均具有重要意義。腫瘤、許多遺傳性疾病、某些神經系統疾病等的發生均與細胞骨架的異常有關。臨床上,常利用細胞骨架在不同細胞內的特異性分布的特徵,來診斷某些疑難疾病,並依據細胞骨架與疾病的關係來設計並指導用藥,開展防病治病。
細胞在病理情況下常常會出現細胞骨架系統異常。如阿爾茨海默症患者,在腦神經元中發現有大量扭曲變形的微管和大量受損的中間纖維;在惡性轉化的細胞中,常表現為微管減少和解聚,細胞骨架異常可增強癌細胞的運動能力。研究表明,微絲束及其末端黏着斑的破壞以及肌動蛋白小體的出現,與腫瘤細胞的浸潤和轉移特性有關。
此外,中間纖維的分布具有嚴格的組織特異性,絕大多數腫瘤細胞在發生轉移後仍表現其原發腫瘤的中間纖維類型,故可作為臨床腫瘤的鑑別診斷和腫瘤細胞是否轉移的判據。中間纖維顯微技術與羊膜刺穿結合,可用於先天胎兒畸形的診斷,例如,若羊水中含有神經元纖維和神經膠質纖維細胞,則提示胎兒或有中樞神經系統畸形。
腫瘤
(一)細胞骨架在腫瘤細胞中的變化
機體中各組織細胞的結構和功能是密切相關的,細胞骨架無論在組裝還是分布上若發生了變化,必將影響到細胞的功能。在惡性轉化的細胞中,常表現為細胞骨架結構的破壞、組裝和分布的異常、微管的解聚等。
我國學者對胃癌、鼻咽癌、食管癌、肺鱗癌、肺小細胞癌、肺腺癌、小鼠肉瘤等9株腫瘤細胞進行觀察,發現腫瘤細胞質內免疫熒光染色的微管減少甚至缺如,且癌細胞的微管變化主要發生在間期,而在分裂期,紡錘體微管與正常細胞相同。用熒光抗體技術已證明,在長期傳代的癌變細胞內微管顯著減少。因此,微管數量的減少是細胞惡性轉化的重要標誌。而且腫瘤細胞內原有的微絲束明顯減少甚至消失,常出現肌動蛋白凝聚小體;腫瘤細胞內的微絲分布異常,無序紊亂,常不與細胞膜相連。
腫瘤細胞的浸潤轉移是一個極其複雜的過程,在此過程中,某些細胞骨架成分的改變可增強癌細胞的運動能力。研究表明,微絲束和其末端黏着斑的破壞以及肌動蛋白小體的出現,可能與腫瘤浸潤轉移的特性有關,肌動蛋白小體形成,可能代表腫瘤細胞高轉移的惡性表型。
(二)中間纖維與腫瘤診斷
中間纖維形態相似,但具有嚴格的組織特異性,不同類型的中間纖維嚴格地分布在不同類型的細胞中,故可根據中間纖維的種類區分上皮細胞、肌肉細胞、間質細胞、膠質細胞和神經細胞。在這幾種纖維類型的基礎上,中間纖維還可進一步分出若干亞型。因絕大多數腫瘤細胞在生長時,繼續保持其來源細胞的中間纖維的種類、超微結構和免疫學特性,如癌是以上皮細胞的角質蛋白為特徵的,肌肉瘤是以結蛋白、非肌肉瘤是以波形纖維蛋白、神經膠質瘤是以神經膠質纖維酸性蛋白、從交感神經來的腫瘤是以神經纖維蛋白(necotinamide protein)為特徵的。所以,人們可根據中間纖維的種類,來鑑別、區分不同組織來源的腫瘤細胞及各腫瘤細胞的亞型,為腫瘤的診斷和治療提供決定性的依據。
在臨床上,中間纖維蛋白的熒光素標記抗體技術在細胞分類尤其是腫瘤細胞的鑑別上,具有廣泛的應用前景。如採用角蛋白熒光素標記抗體確認上皮癌,用波形蛋白熒光素標記抗體確認淋巴肉瘤、黑色素瘤、骨肉瘤等,用結蛋白熒光素標記抗體確認肌細胞肉瘤,用神經纖維蛋白熒光素標記抗體確認神經母細胞瘤、神經節母細胞瘤、嗜鉻細胞瘤等。
(三)微管和微絲與抗腫瘤藥物
在有絲分裂的中/後期,秋水仙素和長春花鹼等化合物可與紡錘體微管蛋白或微管結合,抑制細胞增殖。研究表明具有抗有絲分裂能力的藥物如紫杉醇和長春新鹼等抑制細胞增殖和殺死腫瘤細胞的主要機制是穩定紡錘體微管動力學,而不是使微管解聚或過度多聚化,在有絲分裂的中/後期抑制細胞分裂,誘導細胞凋亡。
細胞鬆弛素是真菌的代謝產物,也是研究最多且應用最廣泛的微絲特異性工具藥,它作用於肌動蛋白,可與微絲正端結合,抑制其聚合,使微絲解聚,導致細胞表麵皮質層松解,引起細胞表面起泡,使微絨毛變成茬狀物,細胞整體形態呈現樹枝狀化,並可抑制各種依賴於微絲的運動,具有抗腫瘤潛能。
神經系統疾病
阿茨海默病(Alzheimer’s disease,AD)即早老性痴呆病,屬微管遺傳性疾病。對患者腦脊液分析發現,AD患者腦脊液中tau蛋白含量明顯高於非AD患者和正常人,且其神經元中還可見到大量損傷的神經元纖維,它們由成對的螺旋狀纖維(paired helical filament,PHF)和相對較直的纖維(straight filament,SF)組成,主要成分是高磷酸化狀態的tau蛋白,其性能穩定。對死亡AD患者的大腦進行分析發現,神經元中微管蛋白的數量並無異常,但微管聚集缺陷。孤立的微管蛋白與結合的微管蛋白均可以高磷酸化的方式與其他配體結合形成穩定的tau蛋白。因為微管是軸漿流必需的細胞骨架,因此AD中微管聚集缺陷,可使微管聚合發生障礙,微管扭曲變形,可能引起軸漿流阻塞,導致神經元纖維包涵體的形成,從而使神經信號傳遞紊亂,影響軸質的物質運輸,使神經元的營養和代謝產生障礙,從而出現痴呆現象。
Tau蛋白及其他一些細胞骨架蛋白的異常還可引起其他神經系統疾病,如運動神經元疾病、帕金森病、肌強直性營養不良等。
遺傳性疾病
某些遺傳性疾病常與細胞骨架的異常或細胞骨架蛋白基因的突變有關。WAS (Wiskoff-Aldrich syndrome)是一種遺傳性免疫缺陷疾病,其特徵是濕疹、出血和反覆感染。研究表明,微絲的異常是引起WAS的根源所在。
隨着研究方法和手段的不斷改進,尤其是利用轉基因小鼠或基因敲除小鼠進行研究,發現中間纖維與許多遺傳疾病有關。人類遺傳性皮膚病單純性大皰性表皮鬆解症(epidermolysis bullosasimplex,EBS)是最典型的例證,該病是由角蛋白14(CK14)基因突變所致。
衰老
老年病學研究表明,老年人隨着年齡的增加,機體各細胞均出現功能低下的表現。這與細胞骨架的數量、結構及功能的變化有關。動物實驗表明,老齡動物的神經元內微管數量減少,腹腔巨噬細胞內的微絲數量減少,可影響神經信號傳遞,影響軸質的物質運輸,影響神經元的營養和代謝,影響免疫機能,進而影響到細胞的功能。所以,老年人的衰老表現為腦功能衰退和機體免疫等多系統多功能的低下。
其他蛋白
不僅如此,細胞骨架還包含有很多結構單元的附屬蛋白質,比如:
分子馬達(molecular motors): 動力蛋白(dynein), kinesin, myosin 結合蛋白:vinculin, cofilin, tropomyosin等等
廣義的細胞骨架還包括核基質(nucleoskeleton)、核纖層(nuclear lamina)和細胞外基質(extracellular matrix),形成貫穿於細胞核、細胞質、細胞外的一體化網絡結構。
發現歷史
細胞骨架(cytoskeleton)是指真核細胞中的蛋白纖維網絡結構。發現較晚,主要是因為一般電鏡制樣採用低溫(0-4℃)固定,而細胞骨架會在低溫下解聚。
直到20世紀60年代後,電鏡制樣採用戊二醛進行常溫固定,人們才逐漸認識到細胞骨架的客觀存在。真核細胞藉以維持其基本形態的重要結構,被形象地稱為細胞骨架,它通常也被認為是廣義上細胞器的一種。細胞骨架不僅在維持細胞形態,承受外力、保持細胞內部結構的有序性方面起重要作用,而且還參與許多重要的生命活動,如:在細胞分裂中細胞骨架牽引染色體分離,在細胞物質運輸中,各類小泡和細胞器可沿着細胞骨架定向轉運;在肌肉細胞中,細胞骨架和它的結合蛋白組成動力系統;在白細胞(白血球)的遷移、精子的遊動、神經細胞軸突和樹突的伸展等方面都與細胞骨架有關。另外,在植物細胞中細胞骨架指導細胞壁的合成。
參考文獻
- ↑ 細胞生物學 | 細胞骨架_蛋白質_纖維_中心搜狐網
- ↑ 什麼是細胞骨架?-資訊分析測試百科網wiki版