间隙连接查看源代码讨论查看历史
| 间隙连接 |

{kind=link}
间隙连接也可以称为连接或黄斑通信 。
(虽然大多数神经组织没有间隙连接,但是当在神经元或神经中发现时,它也可以被称为电突触 ,如牙髓中的神经细胞。)虽然ephapse与间隙连接有一些相似之处,但是通过现代定义,两个是不同的。
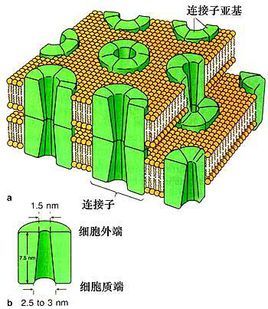
间隙连接是多种动物细胞类型之间的特殊细胞间连接。 它们直接连接两个细胞的细胞质 ,是各种分子 , 离子和电脉冲直接通过细胞之间的调节门。
一个间隙连接通道由两个连接子 (或半通道)组成,它们跨越细胞间隙连接。
间隙连接类似于连接植物细胞的胞间连丝 。
除了成人完全发育的骨骼肌和移动细胞类型(如精子或红细胞)之外,几乎在身体的所有组织中都存在间隙连接。 然而,在诸如海绵和粘液霉菌之类的较简单的生物体中没有发现间隙连接。
基本信息
中文名; 间隙连接
含义; 连接子进行的细胞间连接
性质; 科学
类别; 生物
结构
在脊椎动物中 ,间隙连接半通道主要是连接 蛋白的同源或异源 六聚体 。 无脊椎动物间隙连接包含来自innexin 家族的 蛋白质 。 Innexin与连接蛋白没有显着的序列同源性 。 虽然序列与连接蛋白的序列不同,但innexins与连接蛋白相似,表明innexins以与连接蛋白相同的方式在体内形成间隙连接。 最近表征的pannexin家族最初被认为形成细胞间通道(具有与innexins相似的氨基酸序列 ),实际上起到单膜的作用与细胞外环境相通的通道,已被证明可以通过钙和ATP 。
在间隙连接处,细胞间隙在2到4nm之间并且每个细胞膜中的单位连接子彼此对齐。
由两个相同的半通道形成的间隙连接通道称为同型,而具有不同半通道的那些是异型的。 反过来,均匀连接蛋白组成的半通道被称为同源,而具有不同连接蛋白的那些是异聚的 。 通道组成被认为影响间隙连接通道的功能。
在对innexins和pannexin进行了充分表征之前,基于基因定位和序列相似性 ,将编码连接蛋白间隙连接通道的基因分为三组之一:A,B和C(例如, GJA1 , GJC1 )。 然而,连接蛋白基因不直接编码间隙连接通道的表达; 基因只能产生构成间隙连接通道的蛋白质。
组织层次
DNA转化为连接蛋白的RNA。
一种连接蛋白具有四个跨膜 结构域6 Connexins创建一个Connexon(半通道)。 当不同的连接蛋白连接在一起形成一个连接子时,它被称为异聚连接子 跨细胞膜连接在一起的两个半通道包括间隙连接通道。 当两个相同的连接子聚集在一起形成间隙连接通道时,它被称为同型GJ通道。
当一个同聚连接子和一个异聚连接子聚集在一起时,它被称为异型间隙连接通道。 当两个异聚连接子连接时,它也称为异型间隙连接通道。 几个间隙连接通道(数百个)组装在称为间隙连接斑块的大分子复合物中。
连接通道对的属性
允许细胞之间的直接电通信,尽管不同的连接蛋白亚基可以赋予不同的单通道电导,从约30pS到500pS。
允许细胞之间的化学通信,通过传输小的第二信使 ,如肌醇三磷酸 ( IP 3 )和钙( Ca ), 虽然不同的连接蛋白亚基可以赋予特定小分子不同的选择性。 一般而言,允许小于485 道尔顿 (通过无脊椎间隙连接 1,1 1,100道尔顿)的分子的跨膜运动,尽管不同的连接蛋白亚基可以赋予不同的孔径和不同的电荷选择性。 大的生物分子,例如核酸和蛋白质 ,通过间隙连接连接蛋白通道被排除在细胞之间的细胞质转移中。确保通过间隙连接的分子和电流不会泄漏到细胞间隙中。迄今为止,五种不同的功能归因于间隙连接蛋白:
细胞之间的电和代谢耦合通过半通道进行电和代谢交换肿瘤抑制基因( Cx43 , Cx32和Cx36 )粘附功能独立于导电间隙连接通道(新皮层中的神经迁移) 羧基末端在信号传导细胞质通路中的作用(Cx43)
发生和分配
已经在细胞彼此接触的各种动物器官和组织中观察到间隙连接。 从20世纪50年代到70年代,他们在小龙虾神经, 大鼠胰腺,肝脏,肾上腺皮质,附睾,十二指肠,肌肉, 水蚤 肝脏 , Hydra肌肉, 猴子视网膜, 兔角膜, 鱼胚盘 , 青蛙胚胎, 兔子卵巢, 再聚集细胞, 蟑螂血细胞胶囊, 兔皮, 小鸡胚胎, 朗格汉斯人类胰岛, 金鱼和仓鼠压力传感声前庭受体, 七鳃鳗和被囊心脏, 大鼠曲细精管, 子宫肌层 , 眼镜片和头足类消化上皮。 自20世纪70年代以来,在几乎所有相互接触的动物细胞中都发现了间隙连接。 到20世纪90年代,共聚焦显微镜等新技术可以更快速地检测大面积组织。 自20世纪70年代以来,甚至传统上被认为可能具有分离的细胞(例如骨)的组织显示细胞仍然与间隙连接相连,但是很有效。 间隙连接似乎存在于所有动物器官和组织中,除了通常不与相邻细胞接触的细胞外,发现其他例外将是有趣的。 成人骨骼肌是一个可能的例外。 可以认为,如果存在于骨骼肌中,间隙连接可能在构成肌肉的细胞之间以任意方式传播收缩。 至少在某些情况下,可能不是如其他具有间隙连接的肌肉类型所示的情况。 通过分析癌症 或衰老过程可以指示间隙连接减少或缺失的结果。
功能
可以看到间隙连接在最简单的水平上起作用,作为电流,小分子和离子的直接细胞到细胞途径。 如下所述,对这种通信的控制允许对多细胞生物的复杂下游效应。
胚胎,器官和组织发育
在20世纪80年代,已经研究了间隙连接通信的更微妙但同样重要的作用。 发现通过将抗连接蛋白抗体添加到胚胎细胞中可以破坏间隙连接通讯。 具有阻塞间隙连接区域的胚胎未能正常发育。 抗体阻断间隙连接的机制尚不清楚,但进行了系统研究以阐明其机制。 这些研究的细化表明,间隙连接似乎是细胞极性发展的关键和动物的左/右对称/不对称。 虽然确定身体器官位置的信号似乎依赖于间隙连接,因此在胚胎发育的后期阶段细胞的基本分化也是如此。 还发现间隙连接是造成药物产生影响的信号传递的原因 ,相反,一些药物被证明可以阻断间隙连接通道。
差距交界处和"旁观者效应
细胞死亡
"旁观者效应"及其无辜旁观者被杀的内涵也是由间隙连接介导的。 当细胞由于疾病或损伤而受损并且开始死亡时,消息通过间隙连接被传递到连接到垂死细胞的相邻细胞。 这可能导致其他未受影响的健康旁观者细胞也死亡。 因此,在患病细胞中考虑旁观者效应非常重要,这为更多资金和研究蓬勃发展开辟了道路。后来,对于受辐射或机械损伤损伤并因此伤口愈合的细胞,也研究了旁观者效应。 疾病似乎也会影响间隙连接在伤口愈合中发挥作用的能力。
虽然由于治疗途径的可能性而倾向于关注疾病中的旁观者效应,但有证据表明在组织的正常发育中存在更重要的作用。 一些细胞及其周围基质的死亡可能是组织达到其最终构型所必需的,并且间隙连接对于该过程也是必不可少的。还有更复杂的研究试图结合我们对间隙连接在伤口愈合和组织发育中的同时作用的理解。
电耦合区域
间隙连接通过电和化学方式将细胞连接在大多数动物的身体上。 电耦合可以相对快速地起作用。本节中的组织具有众所周知的功能,观察到由间隙连接协调,其中细胞间信号发生在微秒或更短的时间帧内。
心脏
缝隙连接在心肌中特别重要:收缩信号有效地通过间隙连接,使心肌细胞一致收缩。除了成人完全发育的骨骼肌和移动细胞类型(如精子或红细胞)之外,间隙连接几乎在身体的所有组织中表达。 几种人类遗传疾病与间隙连接基因的突变有关。 其中许多影响皮肤,因为这种组织严重依赖于间隙连接通讯来调节分化和增殖 。 心脏间隙连接可在药理学上用rotigaptide打开。
神经元
位于神经元中的间隙连接通常被称为电突触 。在描述间隙连接结构之前,使用电测量发现电突触。电突触存在于整个中枢神经系统中,并且已在新皮质 , 海马 , 前庭神经核 , 丘脑网状核 , 蓝斑 , 下橄榄核 , 三叉神经中脑核, 腹侧被盖区 , 嗅球 , 视网膜和脊椎动物的 脊髓 。
对蓝斑和神经 胶质细胞与Bergmann神经胶质细胞之间的小脑神经 胶质细胞偶联有一些弱神经元的观察。似乎星形胶质细胞通过间隙连接与其他星形胶质细胞和少突胶质 细胞偶联。此外,间隙连接基因Cx43和Cx56.6中的突变导致白质变性,类似于在Pelizaeus-Merzbacher病和多发性硬化中观察到的。
在神经元间隙连接处表达的连接蛋白包括:m CX36 mCX5 m CX45用mRNA检测至少五种其他连接蛋白(m Cx26 ,m Cx30.2 ,m Cx32 ,m Cx43 ,m Cx47 )但没有超微结构限定的间隙连接中相应蛋白的免疫细胞化学证据。那些mRNA似乎被细胞类型和细胞谱系特异性的微干扰RNA( miRNA )下调或破坏。
视网膜
视网膜内的神经元在一种细胞类型的群体内和不同的细胞类型之间显示出广泛的偶联。[1]
参考文献
- ↑ 间隙连接的结构和功能,豆丁网 , 2014-01-20