木质部
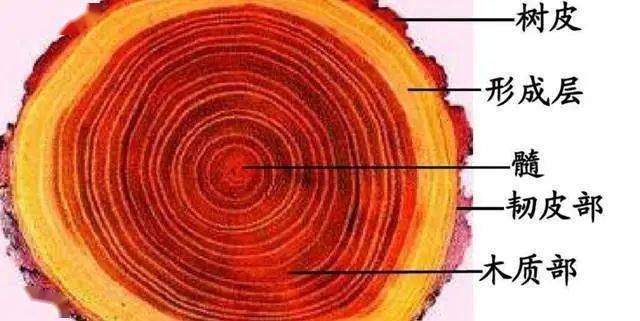 |
木质部是树干的主要组成部分,具有重要的经济价值,[1]在所谓的“硬木”或双子叶木本树种中,木质部由导管、管胞、木纤维和木薄壁组织细胞以及木射线组成,常与韧皮部结合组成维管束,分布在植物体内形成维管系统[2]
目录
定义
木质部指维管植物体内具有输导和机械作用的一种复合组织。由导管、管胞、木纤维和木薄壁细胞等组成,常与韧皮部结合组成维管束,分布在植物体内形成维管系统。导管和管胞为输导水和溶于水的无机盐的管状分子,为木质部的主要组成部分。
分类介绍
导管
导管( vessel)是由多个长管状的死细胞纵向连接而形成的管道,其中每个细胞称为导管分子( vessel member,vessel element)。在导管的形成过程中,每一细胞的横壁逐渐全部溶解而形成单个的大孔隙,或若干个局部溶解而形成数个较小的孔隙。在孔隙处无任何细胞壁的壁层残留,这种孔隙称为穿孔(perforation)。具有穿孔的横壁称为穿孔板( perforation plate)。其中,只具1个穿孔的称单穿孔板,具2个以上穿孔的称复穿孔板。穿孔的形成使导管中导管分子问的邻接壁(即横壁)打通,整个导管成为一个贯通的管道,因此其输水的速度远比管胞为快。此外,导管还可通过其侧壁上无次生壁的部位与相邻的其他细胞进行水分的横向输导。导管的次生壁也像管胞的次生壁那样形成纹理。纹理形成的原因,是次生壁在有些地方铺盖,在另一些地方不铺盖,同时这两种地方成互相间隔或穿插式地分布。根据所形成的纹理之不同,将导管分为下列类型。
1.环纹导管(annular vessel)
其次生壁沿着管状初生壁的内表面,形成多个间隔一定距离的环。其管径较小,存在于植物幼嫩器官中,如玉蜀黍、半夏等植物的幼茎中。
2. 螺纹导管(spiral vessel)
其次生壁从管状初生壁的内表面的一端,成螺旋的带状铺向另一端。其管径与环纹导管的管径差不多或者大一些,也多存在于植物的幼嫩器官中,如莲藕(根茎)和豆芽的茎中。
3. 梯纹导管(scalariform vessel)
其次生壁既横向地形成许多个间隔较短距离的环,又纵向地形成若干条长而直的带,并与前者交连。管壁上无次生壁的地方成为许多个呈扁圆形或长方形的单纹孔,这些单纹孔沿纵轴方向排成若干条整齐的、互不交错的队列。这种导管的直径稍大于上述两种导管的直径,多存在于植物的成长器官中,如葡萄的茎、荆三棱的球茎、香附的块茎中。
4. 网纹导管(reticulate vessel)
其次生壁既作一些横向的条状铺盖,也作一些斜向的条状铺盖,有时还作一些纵向的条状铺盖;这些条状次生壁短的多长的少,相互交连;无次生壁的地方成为一些等大或不等大、同形(但不会都呈圆形)或不同形的坑(单纹孔),它们之间多少呈交错排列,且离得较近。这种导管的管径较大,多存在于器官的成熟部分,如大黄的根及根茎、南瓜的茎和前胡的根中。
5. 孔纹导管(pitted vessel)
其导管壁上无次生壁的地方为一些具缘纹孔或直径较小且呈圆形的单纹孔。其管径较大,多存在于器官的成熟部分,如甘草的根、威灵仙的根和根茎中。
以上所述为5种纹理很典型的导管。在实际的观察中常可以在同一导管上见到呈两种纹理的次生壁,如环纹和螺纹次生壁,梯纹和网纹次生壁等。
某些植物的导管在发育过程中,其邻接的薄壁细胞发生膨胀,进而通过导管壁上无次生壁的部位或纹孔侵入导管腔内,形成大小不等的囊泡状突起。
管胞
管胞( tracheid)是绝大多数蕨类植物和裸子植物的输导组织,同时兼有支持作用。有些被子植物或被子植物的某些器官中也有管胞,但不是主要的输导组织。
管胞中的每个细胞,全形呈两端尖的长管状,成熟后无原生质体,细胞壁含次生壁层且木质化。次生壁形成各种纹理,因而有环纹管胞、螺纹管胞、梯纹管胞、孔纹管胞等类型。管胞的管径较导管的小,且水分从一个管胞细胞运行到另一个管胞细胞的方式,是从纹孔对处渗透过去,因此其运输水分的效率较导管低,是一类原始的输水组织。管胞的主要类型有5种:环纹管胞、螺纹管胞、梯纹管胞、孔纹管胞、纹孔。
木纤维
双子叶木本植物的次生木质部内的纤维。细胞壁厚,细胞腔较窄,形体细长,木质化,是一种厚壁组织,有退化程度不同的具缘纹孔。木纤维有两种主要类型:(1)纤维管胞,具有具缘纹孔;(2)韧型纤维,细胞壁较厚,具单纹孔。木纤维主要起支持作用,有贮藏和输导作用。
木薄壁组织细胞
在次生木质部中一般可分为轴向薄壁组织细胞和径向薄壁组织细胞(即“木射线”)两种。其细胞成熟后,一般都具有加厚的次生壁,并木质化。如有次生壁,则厚壁组织细胞壁之间的纹孔对有具缘、半具缘的类型。木薄壁组织细胞主要起贮藏作用。在木材横切面上,依据轴向薄壁组织细胞与导管相对位置的不同,可分为离管薄壁组织和傍管薄壁组织两大类。
木射线
位于形成层以内次生木质部中的维管射线。其主要功能是横向运输和贮藏。在横切面上,木射线呈辐射状排列。在切向切面上,射线呈纵线或纺锤形。在径向切面上,射线呈不同高度的线状或片状排列。木射线单列或多列、高度从一层到多层细胞。小的射线也可聚合成群,形成聚合射线。组成木射线的薄壁组织细胞,依据细胞最长轴方向的不同,可分为径向伸长的横卧细胞,以及方形或沿纵向伸长的直立细胞两种类型。如果由同一种类型细胞组成的射线,或仅由射线薄壁组织细胞组成的,称为同型射线,由不同类型的薄壁组织细胞组成的射线(双子叶植物)或在裸子植物中由木薄壁组织细胞和射线管胞构成的射线,称异型射线。
因此我们把树干或枝干的木质看做是由庞大的连锁的微观细胞丛组成的,其中有些在器官功能成熟后变得高度木质化并死去;其他与此交织在一起的成分(组成术质部的薄壁组织和放射线)仍然存活,而且代谢活动的程度不同,所有这些细胞来源于精细的分生(周期性分裂)细胞柱体,术语叫做维管形成层,它们形成术质部和树皮之问的分界线毋庸置疑,维管形成层作为一种分生组织或者形成组织,不仅在树的发育过程中发挥重要的作用,而且在树的整个生命历程中都具有重要的作用。
特征
木质部细胞的壁多数木质化,故有一定的机械支持作用。由茎与根的近端部的原形成层分化形成初生木质部,多年生木本植物由于维管形成层的逐年活动,不断增长次生木质部,从而长成粗壮的树干。
木质部解剖学家习惯研究3种薄切面:横切面、径向切面和切向切面。径向切面是通过茎的中心(直径)所作的纵切面,切向切面是垂直于茎的半径所作的纵切面。从微观上看这3种方式结合在一起足以提供鉴别木质部所需要的所有信息:在大多数情况下,可以鉴别出木质所从属的属,有时甚至能鉴别其所属种类。虽然通过该过程我们可以了解木质部许多特点,但是还有其他可利用的特征,不需要显微镜就可以检测到,其中包括一些结构特征用放大镜或者凭肉眼就可以看到,例如颜色和比重,以及树皮的形态。在分析木质部颜色的时候,通常必须考虑活的外层区域和通常没有活性的核心区域的差异——前者一般是颜色较浅的边材,而后者是颜色较深的心材。从商业角度来看,心材是最有价值的,它里面包含有数量不等的树胶、树脂和作为天然防腐剂的沉积物。对于有些木材,这种心材具有与众不同的颜色,例如黑檀树的是黑色的,豆科的洋苏木树的是紫色的。
木材的比重差异很大,正是这一重要的物理特征使得人们可以据此分类:柚木是价值不菲的木材,木质密实、厚重,并且由于里面有树脂材料沉积所以特别不易腐烂;与此相反,美国香脂木特别轻,有点类似于分布比较广泛的西非树,有一定的价值。毋庸置疑,树的用途与显微镜下可观察到的它的解剖结构密不可分。
木质素
在树木生长以及存活的时候,树的导管壁、管胞壁和纤维壁充满一种叫做木质素的物质:当它们死亡后,这种物质仍然存在,因此保留了硬的细胞形状。
木质素是一系列化学成分不详的复合碳化合物的通用术语,但是很明显有许多不同种类的木质素。一个值得一提的有趣事实是针叶树的木质素与阔叶树的木质素不同。另外我们需要注意的重要事实是:这些细胞和它们的木质素决定木质的强度和硬度,因此木质素壁越厚.这些特性就越明显。
生长
维管植物的初生生长和分化时,由原形成层分化的木质部,称为初生木质部,其中含有纤维和薄壁组织细胞,但没有射线,所以,不构成轴向系统和径向系统。在茎、叶和花等部位中,初生木质部和初生韧皮部结合成束状,即为维管束。在茎的维管束之间有薄壁组织,称为髓射线。在根中,初生木质部在中央形成一个核心,核心中央具髓或不具髓。根据分化的早晚,初生木质部又可分为原生木质部和后生木质部两部分。
原生木质部
木质部中最早形成的部分。在茎、叶中,原生木质部是在活跃伸长的组织中成熟的,因此会受到引力的影响,其中无生命的管状分子被拉伸而毁坏。在根中,原生木质部管状分子成熟的地方,是在伸长区以后,所以存在时间较久。原生木质部一般只有管状分子及其包围它们的薄壁组织。原生木质部的管状分子,主要有环纹和螺纹加厚。
后生木质部
初生木质部中,原生木质部分化以后所形成的部分,如有次生木质部时,则在次生木质部发生之前产生。后生木质部一般是在初生植物体仍旧生长时开始发生,其成熟则是在伸长完成以后,因此,比原生木质部受周围组织伸长生长的影响小。后生木质部的组成比原生木质部复杂,除了管状分子和薄壁组织外,还有纤维。薄壁组织可以散布在管状分子之中,也可以形成射线状的径向行列。在切向切面上,它们是轴向的薄壁组织细胞。后生木质部的管状分子可以有环纹、螺纹、梯纹以及具缘纹孔等。
次生木质部
维管植物次生生长时,由维管形成层产生的木质部。次生木质部分为轴向系统和径向或射线系统。轴向系统指次生维管组织中,由纺锤状形成层原始细胞衍生的细胞,它们的长轴与植物体轴呈平行排列。径向系统是次生维管组织中所有射线的总称。其细胞的长轴与体轴间成水平方向排列。
结构
年轮
人们都知道在拦腰劈开树干之后会看到年轮,这种放射状的生长轮代表着它的生长年龄,但是生长年轮是怎样出现的呢?答案就在于维管形成层在每年不同时期产生的细胞类型和大小。在有些例种中,年轮看起来更加明显,这是因为春季初次形成的导管比夏季形成的导管要大得多,英国橡树删岑树部很好地表现这种特征,这类木质称为“环孔材”,有些木材,包括山毛榉树、柳树、苹果树等,在春季和夏季木质之间导管直径没有明显的差异,这类术质的年轮叫做“散孔材”,其年轮问的界线并不十分清晰——夏季这类木质除了导管尺寸外,还存在其他明显的特征作为分界标记。在种软木中,例如苏格兰松,在生长周期结束时形成管胞,此时正是夏季,它们比起春季形成的管胞具有更窄的孔和更厚的壁,由此人们能够识别每年生长的轮。
单子叶的被子植物例如棕榈一般没有显示出由于维管形成层柱所致的每年树干的变粗,也就是说没有明显的次生增厚过程。但是有极少数单子叶被子植物树干变粗的方式非常怪,完全不同于双子叶植物,最著名的例子就是龙血树,它的基本组织里的细胞是分生的―能够像我们前面描述的双子叶植物的形成层细胞那样分裂,但是它不是向外茎部切掉韧皮部细胞,也不是向着中心切断木质部细胞,而是韧皮部细胞和木质部细胞都向着茎的中心被切掉,这样我们会看到如同在初生茎里维管束那样的模式不断复制、增加。
叠生木材和非叠生木材
在种子植物木材中,轴向细胞和射线在切向切面上排列成同一水平方向,称为叠生木材。叠生木材是由叠生形成层所产生。例如,刺槐属的木材。非叠生木材是由非叠生形成层产生的,木材中轴向细胞和射线细胞在切向切面上并不排列成水平系列。例如胡桃属的木材。
边材和心材
根和茎的木材外部,含有生活的细胞和贮存物质、并具输导水分和无机盐功能的部分,称为边材。在次生木质部内部已经停止贮藏和输导作用的部分,称心材,其颜色一般比边材深。边材的数量较为恒定,心材的相对数量在各种植物中都不同,而且还受环境的影响。一般说来,心材的数量可以不断增加,在少数植物中,也可不形成心材。
软材与硬材
软材指裸子植物,特别是松柏类植物的木材。裸子植物木材的结构比较简单,大多数种类最明显的特征是没有导管,主要由管胞组成。在软材中,可以有纤维管胞,但是没有韧型纤维。硬材指双子叶植物的木材。双子叶植物的木材结构比裸子植物的变化大。原始无导管的双子叶植物木材结构比较简单,但在具导管的双子叶植物木材中,还有管胞、一种或几种纤维、轴向薄壁组织细胞和一种或几种射线。另外,在许多双子叶植物木材中,当导管变得不活跃以后,与导管相邻的轴向薄壁组织细胞和射线薄壁组织细胞,可以通过纹孔形成突起,填充在导管腔内,形成侵填体。
散孔材与环孔材
在双子叶植物次生木质部的横切面上,生长轮中的导管管孔分布相当均匀,或从早材到晚材的管孔大小逐渐变化,这种木材称散孔材,例如毛白杨。环孔材是指双子叶植物的次生木质部中,早材的导管管孔明显地比晚材大,因而在横切面上形成清楚的同心[[环状排列。例如水曲柳。
应力木是茎或枝条在抵抗迫使它们倾斜或弯曲的重力作用下形成的木材。松柏类植物和双子叶植物的应力木在产生部位与结构等方面都有相当大的差别,因此又可分为压缩木和伸张木两种。压缩木,即松柏类植物的应力木。在这类植物中,倾斜或弯曲茎或枝条下侧的木材与正常的木材相比,组织质地致密,木质化程度较高,比重大,颜色深,管胞较短。在横切面上,管胞的孔径多为圆形。伸张木,双子叶植物的应力木。在倾斜或向上卷曲茎或枝条上侧的木材,其特点是具胶质纤维,比正常木材木质化程度低,但纤维素含量较高。伸张木也存在于根中。
参考文献
- ↑ 木材知识大全知乎
- ↑ 什么是木质部?什么是木质素?一米养殖