三葉蟲(學名:trilobite)檢視原始碼討論檢視歷史
| 三葉蟲(學名:trilobite) |
 |
三葉蟲(學名:trilobite)是節肢動物門中已經滅絕的三葉蟲綱中的動物。它們最早出現於寒武紀,在古生代早期達到頂峰,此後逐漸減少至滅絕。最晚的三葉蟲於二億五千萬年前二疊紀結束時的生物集群滅絕中消失。三葉蟲是非常知名的化石動物,其知名度可能僅次於恐龍。在所有的化石動物中三葉蟲是種類最豐富的,至今已經確定的有九(或者十)個目,一萬五千多個物種。大多數三葉蟲是比較簡單的、小的海生動物,它們在海底爬行,通過過濾泥沙來吸取營養。它們身體分節,有帶溝將身體分為三個垂直的葉。
基本信息
中文名; 三葉蟲
門; 無脊椎動物門
綱; 三葉蟲綱
目; 球接子目、萊得利基蟲目、
分布區域; 在英屬哥倫比亞、紐約州、中國、德國
基本簡介
三葉蟲是距今5.6億年前的寒武紀就出現的最有代表性的遠古動物,5億~4.3億年前發展到高峰,至2.4億年前的二疊紀完全滅絕,前後在地球上生存了3.2億多年,可見這是一類生命力極強的生物。在漫長的時間長河中,它們演化出繁多的種類,有的長達70厘米,有的只有2毫米。背甲為兩條背溝縱向分為一個軸葉和兩個肋葉,因此名為三葉蟲。
介紹
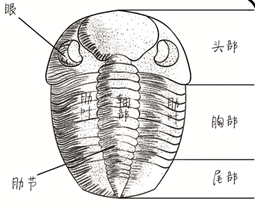
從背部看去三葉蟲(trilobites)為卵形或橢圓形, 成蟲的長為3~10厘米,寬為1~3厘米。小型的6毫米以下 。從結構上可分為頭甲、胸甲和尾甲三部分。三葉蟲體外包有一層外殼,堅硬的外殼為背殼及其向腹面延伸的腹部邊緣 。腹面的節肢為幾丁質,其他部分都被柔軟的薄膜所掩蓋。一般所採到的三葉蟲化石都是背殼。三葉蟲背殼的中間部分稱為軸部或中軸,左、右兩側稱為肋葉或肋部。三葉蟲殼面光滑。或有陷孔、瘤包、斑點、放射形線紋、同心圓線紋、短刺等。頭部多數被兩條背溝縱分為三葉,中間隆起的部分為頭鞍及頸環 ,兩側為頰部,眼位於頰部。頰部為面線所穿過,兩面線之間的內側部分統稱為頭蓋,兩側部分稱為活動頰或自由頰。胸部由若干胸節組成 ,形狀不一,成蟲2~40節 。中間部分為中軸,兩側稱為肋部。每個肋節上具肋溝,兩肋節間為間肋溝。尾部是由若干體節互相融合而形成的 ,1~30節以上不等。形狀一般半圓形,但變化很大,可分為一中軸和兩肋部。肋部分節,有肋溝和間肋溝。肋部可具邊緣 ,邊緣上亦常有邊緣刺。
三葉蟲腹面的節肢極少保存為化石,迄今為止全世界已發現節肢化石的只有19個種。從奧陶紀到泥盆紀末的一些三葉蟲(比如裂肋三葉蟲目)進化出了非常巧妙的脊椎似的結構。在摩洛哥就發現了這樣的化石。此外在俄羅斯西部、美國俄克拉何馬州以及加拿大安大略省也有帶脊椎結構的化石被發現。這種脊椎結構可能是對於魚的出現的一種抵抗反應。
多數三葉蟲有眼睛,它們還有可能用來作味覺和嗅覺器官的觸角,觸鬚可達20至30厘米。三葉蟲 的眼睛是由方解石(碳酸鈣,CaCO₃)組成的。結晶良好的純淨方解石(冰洲石)是透明的,有些三葉蟲使用單晶的、透明的方解石來組成其每隻眼睛的透鏡。這與大多數其它節肢動物不同,球差,同時提供極好的視覺效果。
典型的三葉蟲眼睛是複眼,每個透鏡為一個拉長的稜鏡。
分類 三葉蟲為雌雄異體,卵生,個體發育過程中經過多次周期性蛻殼,在個體發育過程中,形態變化很大。一般劃分為3個階段:幼蟲期、分節期 、成蟲期。三葉蟲綱可以分為10目 :球接子目、萊得利基蟲目、聳棒頭蟲目、褶頰蟲目、鏡眼蟲目、裂肋蟲目、櫛蟲目、鐮蟲目、砑頭蟲目及齒肋蟲目。
習性 三葉蟲與珊瑚、海百合、腕足動物、頭足動物等動物共生。大多適應於淺海底棲爬行或以半游泳生活,還有一些在遠洋中游泳或遠洋中漂浮生活。生活習性的不同決定着其身體構造不同。底棲三葉蟲身體扁平,有的三葉蟲可鑽入泥沙生活,其頭部結構堅硬,前緣形似扁鏟,便於挖掘。有的頭甲癒合,肋刺發育,尾小,具尖末刺,用以在泥沙中推進。另外,適於在鬆軟或淤泥海底爬行生活的類型,其肋刺和尾刺均很發達,使身體不易陷入泥中。營漂浮生活的類型,往往身體長滿纖細的長刺。它們以原生動物、海綿動物、腔腸動物、腕足動物的動物屍體或以海藻等細小生物為食。
三葉蟲的生活習性是多種多樣的,化石中最多的一類是保存在石灰岩或頁岩中,可見當時它們大多生活在淺海底或游移於淤泥之上。它們有的稍能游泳,有的隨水漂流。志留紀中期的齒蟲類,整個身體幾乎被密密的長刺包圍,這些長刺對於它們在水裡游泳來說是一種強有力的推進器,因此可以推測它們是游泳的能手;同時,這些長刺也是抵禦天敵的有效武器。這種類型的三葉蟲主要是出現於奧陶紀到泥盆紀時期,當時與它共生的鸚鵡螺類、板足鱟類和魚類都是三葉蟲的勁敵,如果三葉蟲不增強它的游泳能力和禦敵的武器,它們怎樣在那個競爭激烈的環境中繼續生存繁衍呢?
奧陶紀的某些三葉蟲,如寶石蟲、斜視蟲、隱頭蟲等還發展了捲曲的能力,它們的頭部和尾部可以完全緊接在一起,僅將背部的硬殼暴露在外;它們還可以鑽進淤泥以保護其柔軟的腹部器官,這樣,一方面便於禦敵;另一方面也可以以類似於尺蠖那樣的伸曲的方式推動身體前進。
對於三葉蟲的個體發育過程,科學家通常是通過採集同一層位中同一種個體的不同生長階段的標本來研究的。三葉蟲的個體發育,大致劃分為三個時期--幼年期、中年期和成年期。幼年期蟲體頭部和尾部尚不分明,也沒有胸節,直徑大約為0.24~1.3毫米。中年期蟲體頭部和尾部已經分開,胸節也已經發育,但是節數比成年期少一節。成年期蟲體的胸部與尾部節數增加到了極限,蟲體增大,殼上的刺、瘤等附加物均出現了。
孵化 從卵中孵化出來的幼蟲被稱為原甲期(protaspid period),在這個階段里所有環全部融合在一起形成一個單一的盔甲。在此後的生長期里在每次蛻皮時在尾部前會增加新的胸部環。此後在蛻皮時環的數目不再增加,對三葉蟲的幼蟲階段人們的認識很豐富,它們為研究三葉蟲之間的親緣關係提供了非常重要的幫助。
起源 三葉蟲的祖先可能是類似於節肢動物的動物,如斯普里格蠕蟲或其它隱生宙埃迪卡拉紀時期類似三葉蟲的動物。早期三葉蟲與伯吉斯頁岩和其它寒武紀的節肢動物化石有許多類似的地方,因此三葉蟲與其它節肢動物可能在埃迪卡拉紀和寒武紀的交界之前有共同的祖先。
演化 三葉蟲發展迅速,在寒武紀晚期達到繁殖高點的時代。為了適應不同的生活環境,形態演變多種多樣。有的頭、胸、尾三部分大小相等,殼體緩平,頭、尾都缺少明顯的裝飾,如大頭蟲 (BUMASTUS):有的頭部既寬且大,前緣被一條平闊的圍邊所環繞,其上還排列着整齊的瘤粒,如隱三瘤蟲(CRYPOLITHUS);有的為了免於受害,在胸、尾裝飾着尖長的針刺,如裂肋蟲(LICHAS);有的殼體還能夠捲曲成為球狀,如隱頭蟲(CALYMENE)。
滅絕 三葉蟲滅絕的具體原因不明,但是志留紀和泥盆紀時期兩齶強大,互相之間由關節連接的鯊魚和其它早期魚類的出現與同時出現的三葉蟲數量的減少似乎有關。三葉蟲為這些新動物可能提供了豐富的食物。
此外到二疊紀後期時三葉蟲的數量和種類已經相當少了,這無疑為它們在二疊紀--三疊紀滅絕事件中滅絕提供了條件。此前的奧陶紀--志留紀滅絕事件雖然沒有後來的二疊紀--三疊紀滅絕事件那麼嚴重,但是也已經大大地減少了三葉蟲的多樣性。
對於三葉蟲的個體發育過程,科學家通常是通過採集同一層位中同一種個體的不同生長階段的標本來研究的。三葉蟲的個體發育,大致劃分為三個時期--幼年期、中年期和成年期。幼年期蟲體頭部和尾部尚不分明,也沒有胸節,直徑大約為0.24-1.3毫米。中年期蟲體頭部和尾部已經分開,胸節也已經發育,但是節數比成年期少一節。成年期蟲體的胸部與尾部節數增加到了極限,蟲體增大,殼上的刺、瘤等附加物均出現了。
化石
形成環境在遠古海洋中三葉蟲的生活環境從淺海到深海非常廣。偶爾三葉蟲在海底爬行時留下的足跡也被化石化了。幾乎在所有今天的大陸上均有三葉蟲的化石被發現,它們似乎在所有遠古海洋中均有生存。
今天在全世界發現的三葉蟲化石可以分上萬種,由於三葉蟲的發展非常快,因此它們非常適合被用作標準化石。在英屬哥倫比亞、紐約州、中國、德國和其它一些地方發現過非常稀有的、帶有軟的身體部位如足、鰓和觸角的三葉蟲化石。
品種 中國三葉蟲化石是早古生代的重要化石之一,是劃分和對比寒武紀地層的重要依據。
主要的三葉蟲化石品種有:蝙蝠蟲(Drepanura)、四川蟲(Szechuanella)、副四川蟲(Parasxechuanella)、似櫛殼蟲(即湘西蟲)、王冠蟲(Coronocephalus)、溝通蟲(Ductina)
三葉蟲的世界
命名 早在300多年前的明朝崇禎年間,一個名叫張華東的人在山東泰安大汶口發現了一種包埋在石頭裡的"怪物",其外形容貌頗似蝙蝠展翅,於是他就為之命名為"蝙蝠石"。到了20世紀20年代,我國的古生物學家對"蝙蝠石"進行了科學研究,終於弄清楚了原來這是一種三葉蟲的尾部。這種三葉蟲生活在5億年前的寒武紀晚期,是海洋中的一種節肢動物。為了紀念這個世界上給三葉蟲起的第一個名字,我國科學家就把這種三葉蟲由拉丁名翻譯成的中文名字依然叫做蝙蝠蟲。 國外研究三葉蟲的最早記錄可以追溯到1698年。當時,魯德把一個頭部長有三個圓瘤的三葉蟲化石命名為"三瘤蟲"。到了1771年,瓦爾其根據這種動物的形態特徵,即身體從縱橫兩方面來看都可以分成三部分:縱向上分為頭部、胸部和尾部,橫向上分為中軸及其兩邊的側葉部分,因而給出了一個恰如其分的名稱--"三葉蟲"。
繁殖 三葉蟲最早是隨着寒武紀初期的小殼動物群而出現的,小殼動物群主要是指軟舌螺、腹足類、單板類、喙殼類和分類位置不明的一大批個體微小(一般僅1~2mm)、低等的軟體動物,當時的海洋條件已經適合於它們生存,這些動物給三葉蟲帶來了豐富的食源,在那時的海洋中,三葉蟲還沒有遇到有力的競爭對手,因此它們橫行霸道,迅速發展,整個寒武紀成了三葉蟲的世界。
身體 三葉蟲的樣子奇特,身體分為頭、胸和腹三個部分。貝殼則有三個葉體,二葉位於縱向軸葉的每一側,因此被稱為:"三葉蟲"。從背部看去三葉蟲微卵形或橢圓形,成蟲的長為3~10厘米,寬為1~3厘米。外殼堅硬,正中突起,兩肋低平,也形成縱列的三部分,三葉蟲的名字就是這麼來的吧。由於三葉蟲的背殼堅硬,所以容易被保存成為化石。我們今天了解這種絕滅了的動物,全是通過化石來認識它們的。三葉蟲的頭部由於覆蓋有硬甲,可稱為頭甲,頭甲上中央隆起的部分叫頭鞍,頭鞍的形狀和大小在不同種類中變化較大,頭鞍前部是頭蓋,上面發育着眼脊、眼葉和眼。頭蓋兩側的邊緣下凹並延展形成活動頰,活動頰常常進一步形成十分尖銳的頰刺,伸向身體的後方,整個頭甲是三葉蟲分類和種屬鑑定的重要依據。
胸甲尾部 胸甲由許多形狀相似的胸節組成,這些胸節相互銜接,與絕大多數節肢動物的體節相似,胸節可以活動,並有彎曲的功能。三葉蟲身體能夠蜷起或伸展開全靠這些活動的胸節,但幼年體的三葉蟲沒有胸節。尾甲是指三葉蟲身體末端由若干體節融合而成的部分,它們形成三葉蟲獨特的尾部。三葉蟲的尾一般都是半圓形,由於尾的邊緣常常形成大小不同的尾刺,使許多三葉蟲的尾伸展、放射,變得很美麗。整個三葉蟲的背面硬而光滑,但科學家們發現有些種類在背甲上具有小瘤或小結節,這些小瘤和小結節與背甲上的頰刺、肋刺、尾刺一起,構成了複雜的防護"盔甲"。
發育 經過各國古生物學家多年的研究,認為三葉蟲具有複雜的發育階段。三葉蟲為雌雄異體,卵生,在它們在一生的發育中,要經過多次的蛻殼才能長成,當前的許多節肢動物都承襲了三葉蟲的生長方式。三葉蟲從幼蟲到成蟲,一般經歷三個生長階段,即幼年期、分節期和成蟲期。了解這點,對我們在野外採集三葉蟲化石很有必要,如果人們稍微具備一些有關三葉蟲發育階段的知識,就能對採集到的三葉蟲化石做出大致的鑑定,不致於把不同發育階段的同一種三葉蟲當做不同形態的屬種了。
幼年期的三葉蟲除身體很小外,常常凸起明顯,頭部與尾部區分不明顯,沒有胸節,蟲體呈圓球狀。以後,隨着三葉蟲不斷生長,胸節逐漸增加,當胸節全部長成不再增加時就進入成年期,此時意味着三葉蟲已達到性成熟階段,能夠生兒育女了。三葉蟲每蛻一次殼,身體都會增大,殼上的刺、瘤、甚至尾甲的分節數也會增加。
三葉蟲長大以後就可以在海洋中無憂無慮地生活了,至今為止,人們還沒有在陸相地層中發現三葉蟲化石,這說明這種動物確實只生存在海洋里。由於三葉蟲化石常常與珊瑚、腕足動物、頭足動物共同出現,表明它們都喜歡生活在比較溫暖的淺海,在那裡,三葉蟲以各種微小的生物為食,或者也對海草及動物的屍體感興趣。可以肯定,它們不具有主動攻擊的能力,因為三葉蟲沒有良好的游泳器官,也不具備流線形的體形,在水中行進的速度較慢。從它們的堅固背甲可以想象,一旦有兇猛的動物(如鸚鵡螺類)向它們擺出進攻的架勢時,三葉蟲會迅速把身體蜷起,象穿山甲那樣把自己保護起來,悄悄沉入海底。
為什麼出現那麼多三葉蟲?
寒武紀時為什麼出現那麼多三葉蟲呢?科學家們通過古生態學的研究認為,三葉蟲具有很好的適應環境的生存方式。三葉蟲並不遵循着單一的生活模式,有些種類的三葉蟲喜歡游泳,有些種類喜歡在水面上漂浮,有些喜歡在海底爬行,還有些習慣於鑽在泥沙中生活,它們占據了不同的生態空間,寒武紀的海洋成了三葉蟲的世界。在寒武紀以後的地質時代,這種不同尋常的生物與其它無脊椎動物又共同生存了很長時間,才逐漸數量減少和衰退。我國三葉蟲化石非常豐富,僅在寒武紀的早期就發現了200多個屬,山東泰安盛產的"燕子石",經研究發現就是當時大量活動的三葉蟲死後堆積形成的,那些顯露在岩石表面紛紛欲飛的"燕子",實際上全是一種長有長長尾刺三葉蟲的尾甲。
三葉蟲出現後,在整個早古生代(包括寒武紀、奧陶紀和志留紀)都可作為眾多生物的代表,它們和許多其它生物一起共同揭開了地球走進生物多樣化的序幕,從此,一個欣欣向榮的生物世界才真正出現。晚古生代時三葉蟲數量隨着門類眾多的海相無脊椎動物的大量湧現而減少,中生代到來時終於絕滅。
摺疊編輯本段動物分類 在動物分類學上,三葉蟲屬於節肢動物門、三葉蟲綱。它們生活在遠古的海洋中,主要出現在寒武紀,到寒武紀晚期時發展到頂點。此後,三葉蟲從極盛的高峰走向衰退,延續到二疊紀末期時絕滅,沒有進入中生代。三葉蟲在整個古生代3億多年的漫長地質歷程中生生不息,繁衍出了眾多的類群和巨大的數量,總計有1500多個屬,1萬多個種,其中發現於我國的有大約500個屬。
三葉蟲的形狀大多為卵圓形或橢圓形,個體大小相差卻很懸殊。發現於葡萄牙奧陶紀地層中的烏拉裂肋蟲是最大的三葉蟲之一,長達70厘米。而古盤蟲、球接子之類的微小三葉蟲卻只有不到6毫米。常見的三葉蟲一般長度都在3到10厘米,寬度在1到3厘米。超過20厘米的就算大型的了。在我國昆明寒武紀早期地層中曾經發現過長度為30厘米的萊得利基蟲。
三葉蟲生活的年代距今雖然遙遠,但是科學家對它的形態、構造等特徵的了解是相當充分的,主要的原因有以下幾點:首先,三葉蟲身體表面披有堅固的甲殼,在個體發育過程中經歷多次脫殼生長,所以它們在地層中遺留下的化石數量比其他生物要多;其次,寒武紀海洋中很少有比三葉蟲更大、更凶暴的動物和它生活在一起,因此它們能夠迅速繁衍,廣泛分布;此外,三葉蟲化石大多保存在質地細緻的石灰岩或頁岩中,因此,不僅外殼的特徵能夠被觀察得很清楚,而且有時其內部構造也能被看得很清晰。
三葉蟲的主要特徵表現在它的背殼構造,其頭部中央有一個突起的"頭鞍",可能是安置腦的處所。頭鞍的表面有的光滑無飾,有的瘤斑點綴,還有的具有為數不等的橫溝。這些橫溝被稱為"頭鞍溝"。頭鞍兩側,一般有成對的眼睛。沿眼睛的前後有一條溝,稱為"面線",這是三葉蟲成長過程中藉以脫殼鑽出身體的地方。頭部腹面的前端有一對分節的觸鬚,既是行動器官,又是感覺器官。觸鬚的後面是攝食的口,通常蓋着"唇瓣"。口兩側有許多細小而分節的行動器官--附肢,附肢上有細密的纖毛,大概可以起到呼吸的作用。
三葉蟲的胸部分節,多者達十幾節,少者只分兩節。各節之間以覆瓦狀(即像房頂的瓦片一樣一片覆疊在另一片的上面)關聯起來,便於捲曲活動。三葉蟲腹面兩側有為數眾多的分節附肢,附肢上具有纖毛,因此這些附肢也兼負行動和呼吸之用。三葉蟲的尾部和胸部一樣,縱向上分為中軸及其兩側的肋葉部,其形態多樣;尾部的邊緣有的帶刺,有的不帶刺。
三葉蟲自從在寒武紀早期出現以後,在整個系統演化中各部主要構造特點也逐漸發生相應的變化,這些變化規律主要有下列幾方面:
1、頭鞍形態的變化:寒武紀早期的原始三葉蟲的頭鞍形態多為長圓錐形,凸起也不顯著。往後到了寒武紀中期以後,頭鞍逐漸縮短,兩側趨向平行,成為圓柱形,甚至有的成為了球形。到了寒武紀晚期及以後的三葉蟲,甚至頭鞍與其兩側的頰部分界也不清楚了。
2、面線後支所在位置的變化:早期三葉蟲的面線後支(即眼睛之後的那段面線)終點常與頭部的後邊緣或兩頰角相交;往後到了奧陶紀以後的類型,則常與頭部的兩旁側緣上相交。
3、眼的變化:某些三葉蟲的眼睛。早期是新月形的,隨後逐漸變小,最後消失。另一類複眼比較發達的三葉蟲,眼睛則由小變大,最後會出現眼柄,眼睛則長在高高聳起的眼柄頂端上。志留紀的許多三葉蟲就屬於這一類。
4、身體周圍長刺的變化:寒武紀和奧陶紀的三葉蟲很少長刺,而志留紀及其以後的類型長刺較為多見,而且刺比以前的也更加複雜。
5、胸節由多變少,尾部由小變大,頭鞍上的橫溝由多到少等等趨勢也在許多類型的三葉蟲中顯示出來。
研究
三葉蟲是節肢動物的一種,全身明顯分為頭、胸、尾三部分,背甲堅硬,被兩條縱向深溝割裂成大致相等的三片,所以叫做三葉蟲。
最早描述的三葉蟲化石在英國,當時誤以為是一種比目魚化石。在十九世紀國外對三葉蟲的研究已經比較透徹,中國的三葉蟲最早由許多國外專家和探險家進行命名和報道,民國時期的盧衍豪教授對中國三葉蟲的研究做了重要的起步工作。新中國建立後,國家開始系統研究中國的三葉蟲。有《中國的三葉蟲》等書出版 。吉林大學博物館館長昝淑芹教授對遼寧東部地區(遼東半島和太子河流域)寒武紀的三葉蟲動物群進行過系統研究。記述了該區寒武紀地層中的四十一個科或亞科,一百零一個屬或亞屬,一百八十九個種的三葉蟲化石,其中有新屬五個,新種57個。發表了專著《遼寧東部寒武紀地層及三葉蟲動物群》等一系列的研究成果。今天在全世界發現的三葉蟲化石可以分為上萬種,由於三葉蟲的發展非常快,因此它們非常適合被用做標準化石,地質學家可以使用它們來確定含有三葉蟲的石頭的年代。三葉蟲是最早的、獲得廣泛吸引力的化石,至今為止每年還有新的物種被發現。但是由於我國三葉蟲分類起步晚,當前我國三葉蟲分類研究方面仍很混亂。
其他
三葉蟲還是中國一種"貢茶"的名稱--三葉蟲茶,城步民間簡稱蟲茶,也可以做茶葉蛋。
感覺器官
許多三葉蟲有眼睛,它們還有可能用來作味覺和嗅覺器官的觸角。有些三葉蟲是瞎的,可能它們居住在非常深的海底,那裡沒有光,因此用不着眼睛。有些(比如蛙形鏡眼蟲Phacops rana)有很大的眼睛。
三葉蟲的眼睛是由方解石(碳酸鈣,CaCO₃)組成的。純的方解石是透明的,有些三葉蟲使用單晶的、透明的方解石來組成其每隻眼睛的透鏡。這與大多數其他節肢動物不同,球差,同時提供極好的景深。在今天生存的動物中蛇尾海星Ophiocoma wendtii使用類似的透鏡。
典型的三葉蟲眼睛是複眼,每個透鏡都是一個一般排列為六邊形的拉長的稜鏡。每隻複眼內的透鏡數不等,有些只有一個,有些可達上千。
郵票上三葉蟲
1958年4月15日我國發行一套《中國古生物》郵票,其中第一枚就是"嵩里山三葉蟲" 自左至右:嵩里山三葉蟲、祿豐龍、腫骨鹿 自左至右:嵩里山三葉蟲、祿豐龍、腫骨鹿[1]